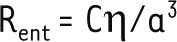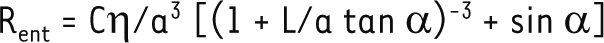
 |
| |||
Med Sci (Paris). 2015 February; 31(2): 174–179. Published online 2015 March 4. doi: 10.1051/medsci/20153102014.Perméabilité optimale des aquaporines Une histoire de forme ? 1Institut lumière matière, UMR5306 université Lyon 1-CNRS, université de Lyon, 3, boulevard du 11 novembre 1918, 69622Villeurbanne, France 2Laboratoire de Physique Statistique, UMR CNRS 8550, École Normale Supérieure, 24, rue Lhomond, 75231Paris Cedex 05, France Corresponding author. | ||||||||
| ||||||||
Les aquaporines forment une famille d’au moins dix protéines transporteuses d’eau. On les trouve aussi bien chez les mammifères que dans les micro-organismes, ou encore les plantes [1–4]. Elles sont situées, entre autres, dans les épithéliums et les endothéliums. Dans un corps humain composé d’au moins 70 % d’eau, l’organisation de l’eau à l’intérieur des compartiments biologiques est fondamentale à la vie. Les aquaporines en sont le « système de plomberie » selon Peter Agre, qui a reçu le prix Nobel de chimie en 2003 pour sa découverte de l’aquaporine [3]. Les cellules doivent en effet absorber, conserver ou excréter les molécules d’eau, sans laisser passer d’autres molécules [5, 6]. La perméabilité de l’aquaporine est ainsi sélective : si les molécules d’eau (de formule chimique H2O) traversent les membranes avec une résistance très faible, ces dernières ne sont en revanche pas perméables à l’ion hydronium (H3O+). Cette propriété est essentielle ; par exemple, si nos reins réabsorbaient cet acide en plus de l’eau, nous développerions des troubles de l’équilibre acido-basique (acidose métabolique). Ce sont les aquaporines qui permettent d’expliquer comment le cerveau sécrète et absorbe le liquide céphalo-rachidien, comment nos yeux génèrent l’humeur aqueuse ou encore comment les reins concentrent l’urée si efficacement. Les performances des aquaporines sont bien supérieures à ce que l’on attendrait d’un « système de plomberie » classique extrapolé aux petites échelles. En d’autres termes, les modèles macroscopiques ne s’appliquent plus pour le canal interne de l’aquaporine de taille nanométrique (un milliardième de mètre, ce qui représente quelques diamètres moléculaires), et de nouveaux comportements d’écoulement spécifiques à l’échelle nanométrique apparaissent [7]. L’exploration de ces comportements est permise par les progrès récents réalisés dans la fabrication de nanosystèmes contrôlés. Un exemple frappant est donné par les mesures de très haute perméabilité des membranes synthétiques constituées de nanotubes de carbone [8, 9]. Dans ce contexte, et dans une perspective de biomimétisme, il est fondamental de comprendre les origines de l’efficacité de certains canaux naturels, tels que les aquaporines. Être capable de réaliser des membranes d’une efficacité comparable permettrait de grandes avancées, aussi bien dans le domaine médical, comme par exemple pour l’amélioration de la dialyse, que pour le dessalement de l’eau de mer, son ultrafiltration ou sa décontamination. Le canal interne des aquaporines se compose de trois parties distinctes : une constriction cylindrique au centre et deux entrées coniques (Figure 1). La constriction centrale est si étroite que les molécules d’eau ne peuvent y circuler qu’en « file indienne », ce qui permet notamment d’assurer la sélectivité des aquaporines. Les aquaporines s’insèrent dans les membranes des cellules et permettent la connexion entre les réservoirs séparés par celles-ci, typiquement les volumes intra- et extracellulaires. L’écoulement entre les réservoirs est généré par une différence de concentration en sel, de part et d’autre de la membrane cellulaire. L’eau a alors tendance à diluer le réservoir le plus concentré ; c’est le phénomène d’osmose. Un exemple de manifestation de l’osmose dans la vie de tous les jours est l’utilisation de solutions salées pour la décongestion des fosses nasales. Le liquide injecté à l’aide d’un spray est dit hypertonique ; il possède une concentration en sel supérieure à la concentration du cytoplasme des cellules. Au contact des cellules de la muqueuse, une force osmotique se créée de part et d’autre de la paroi semiperméable des cellules, entraînant un flux d’eau au travers des canaux, ce qui permet de désengorger ces dernières.
L’étude que nous présentons ici a pour but de comprendre l’efficacité des aquaporines, ces canaux qui permettent une osmose efficace au sein du vivant. C’est un aspect géométrique particulier de ces canaux qui a attiré notre attention : les entrées coniques. Ont-elles un impact sur la perméabilité ? Pour répondre à cette question, nous mettons de côté la chimie complexe qui permet notamment aux molécules d’eau de traverser efficacement la partie centrale de l’aquaporine [11], pour nous focaliser exclusivement sur une étude de l’écoulement dans les zones coniques [12]. | ||||||||
Les écoulements rencontrés dans les aquaporines sont dits rampants, ce qui signifie qu’ils sont gouvernés par la viscosité des liquides (c’est-à-dire leur résistance à l’écoulement), tandis que les effets d’inertie peuvent être négligés. C’est une différence notable avec les écoulements plus couramment rencontrés au quotidien, comme ceux induits dans nos conduits d’alimentation en eau. Ce régime d’écoulement particulier est rencontré dans les écoulements à très faibles vitesses, les écoulements de fluides très visqueux ou encore les écoulements à très petites échelles. La grandeur fondamentale que nous allons utiliser pour quantifier la perméabilité des aquaporines est la résistance hydrodynamique. Pour la définir, nous avons besoin de deux grandeurs. La première est la chute de pression Dp, différence de pression de part et d’autre du canal, qui induit l’écoulement du liquide dans le canal. La seconde grandeur est le débit, noté Q, qui mesure le volume d’eau qui traverse le canal par unité de temps. Le rapport entre la chute de pression Dp et le débit Q définit la résistance hydrodynamique totale du canal : Rtot = Dp/Q. Plus cette résistance est faible, meilleure est la perméabilité. La résistance hydrodynamique est une conséquence globale d’un phénomène local : la dissipation visqueuse. Cette dissipation visqueuse est reliée à une propriété du fluide, sa viscosité de cisaillement h, qui correspond à la contrainte de cisaillement (frottement entre les couches de fluide avançant à des vitesses différentes) qui accompagne un gradient (à savoir, une variation spatiale) de vitesse. Il y a ainsi plus de frottement entre les couches de liquide s’écoulant à des vitesses différentes pour un liquide très visqueux, comme du miel, que pour un liquide moins visqueux, comme le sang. Dans des canaux nanométriques, le frottement du liquide aux parois peut donner lieu à plusieurs régimes d’écoulement. Dans le cas où ce frottement est très fort, la vitesse du liquide s’annule à la paroi : c’est la condition de non-glissement, traditionnellement utilisée pour décrire les écoulements à grande échelle. L’écoulement forme alors un profil de vitesse parabolique dit de Poiseuille (Figure 2A). Le fluide se déplace plus rapidement sur l’axe du tube que près des parois, où sa vitesse est nulle, ce qui donne lieu à de forts gradients de vitesse, donc à une dissipation visqueuse importante. Cependant, de nouveaux outils expérimentaux et numériques pour sonder les écoulements à l’échelle nanométrique, développés durant ces 20 dernières années, ont permis de montrer que les liquides pouvaient glisser sur certaines surfaces qui frottent peu [13]. Pour quantifier cette condition dite de glissement partiel, on introduit une grandeur appelée longueur de glissement. Elle est notée b et représente la distance dans le mur à laquelle il faut extrapoler le profil de vitesse pour atteindre le point de vitesse nulle (Figure 2B). La longueur de glissement est d’autant plus grande que le frottement à la paroi est faible [13]. Ainsi, sur des surfaces qui frottent extrêmement peu, la longueur de glissement devient très grande devant les dimensions du canal. On peut alors faire l’approximation de glissement parfait, qui correspond à une longueur de glissement infinie, ou encore à un frottement strictement nul (Figure 2C). Il en résulte un écoulement dit bouchon, dans lequel le liquide se déplace en bloc dans le canal. Il n’y a donc pas de gradient de vitesse, et la dissipation visqueuse est nulle.
La résistance hydrodynamique interne Rin d’un écoulement rampant dans un tube non glissant est proportionnelle à la longueur du tube L, à la viscosité du fluide η et inversement proportionnelle au rayon du tube a à la puissance 4 :
avec A, un pré-facteur numérique qui dépend de la géométrie du canal. En cas de glissement à la paroi, cette résistance est divisée d’un facteur faisant intervenir la longueur de glissement b : 1+Bb/a, où B est un deuxième pré-facteur numérique tenant compte de la géométrie. À cette résistance interne viennent s’ajouter les effets dus aux connexions entre le tube et les réservoirs. En effet la convergence du liquide à chaque entrée/sortie du canal génère des gradients de vitesse, donc une dissipation visqueuse. Un nouveau terme de résistance, appelé résistance d’entrée Rent, vient donc s’ajouter à la résistance interne du canal [14–16]. Cette résistance est proportionnelle à la viscosité du fluide η, et inversement proportionnelle au rayon du canal a au cube :
avec C, un pré-facteur qui dépend de la géométrie du canal. Dans le cas d’un tube cylindrique, A = π/8, B = 4 et C ≈ 3. Dans le cas d’un canal de longueur L faible devant son rayon a, également appelé pore, ou dans le cas d’un fort glissement à la paroi (longueur de glissement b très grande devant le rayon a), la résistance d’entrée prédomine sur la résistance interne et gouverne, de ce fait, la perméabilité globale du canal. Une estimation rapide montre que, dans les aquaporines, la résistance d’entrée joue un rôle très important : en effet, l’ordre de grandeur typique donné par l’équation 2 est Rent ≈ 1 Pa.s.nm-3, ce qui est très proche des résistances (totales) d’aquaporines trouvées dans la littérature, qui varient typiquement entre 1 et 3 Pa.s.nm-3 [17]. | ||||||||
On considère une membrane séparant deux réservoirs et percée d’un pore, composé d’un canal central et de deux entrées coniques (Figure 1C). Afin de nous focaliser sur le rôle des entrées, nous utilisons un cylindre central parfaitement glissant, dans lequel la dissipation visqueuse sera nulle. La méthode numérique utilisée est appelée méthode des éléments finis. Elle consiste en un découpage de l’espace en éléments de petites tailles, puis en une résolution des équations de la physique continue dans chaque élément, en respectant les conditions de bord appliquées. On applique une différence de pression de part et d’autre de la membrane, ce qui crée un écoulement de fluide au travers du pore. On mesure ensuite le débit, ce qui nous permet de calculer la résistance hydrodynamique en fonction des différents paramètres du problème, comme la longueur des cônes L, l’angle d’ouverture α, ou encore la longueur de glissement sur les parois des cônes b. Dans un premier temps, nous négligeons les frottements aux parois des cônes, en considérant des cônes parfaitement glissants. Nos calculs montrent alors que la résistance du pore possède un minimum pour un angle d’ouverture spécifique, noté α0 (Figure 3). Cet angle optimal décroît lorsque la longueur du cône augmente, et reste globalement compris dans une gamme allant de 5° à 25°.
Afin de comprendre ces observations, on trace le profil de dissipation visqueuse (Figure 4). Lorsque le cône est fermé, la dissipation provient exclusivement des connexions entre les réservoirs et le canal. À mesure que le cône s’ouvre, cette dissipation est transférée progressivement vers les zones de connexion entre les cônes et le tube central. À partir de cette observation, nous allons construire un modèle simple pour la résistance des entrées coniques. Nous faisons une approximation de la résistance due à la connexion entre le réservoir et le cône à l’aide d’une expression classique de type résistance d’entrée (équation 2), en prenant le rayon d’entrée du cône, donné par a + Ltan α.
Pour la connexion entre le cône et le tube central, nous trouvons, en utilisant les calculs numériques, qu’une expression classique de résistance d’entrée multipliée par le sinus de l’angle prédit très bien le comportement observé. On arrive donc, pour la résistance d’entrée totale du canal biconique, à l’expression suivante :
Le premier terme, qui correspond à la contribution de la connexion cône-réservoir, décroît très rapidement avec α, tandis que le second, qui correspond à la contribution de la connexion cône-cylindre, croît beaucoup plus lentement avec α, ce qui explique que l’angle qui minimise la résistance totale est très faible. L’équation 3 reproduit bien les résultats numériques et, en particulier, la dépendance de l’angle optimal avec la longueur du cône (Figure 3B). Nous avons ensuite étendu notre description en considérant un glissement partiel le long des parois des cônes plutôt qu’un glissement parfait. Tant les calculs numériques que le modèle analytique montrent que l’angle optimal augmente lorsque le glissement diminue (Figure 5A).
Revenons à présent à l’aquaporine, qui constituait notre point de départ. Nous comparons les géométries optimales prédites dans le cadre de notre modèle avec les géométries réelles des aquaporines. Pour cela, nous avons extrait des profils obtenus par rayons X publiés dans la littérature, les valeurs des paramètres correspondant à notre modèle (Figure 1B). Il y a trois paramètres géométriques qui nous intéressent : le rayon a de la partie centrale, la longueur du cône L, ainsi que l’angle d’ouverture α. La plupart des aquaporines étant asymétriques, chaque profil donne deux valeurs pour L et α. On compare les valeurs des angles observés dans différentes aquaporines aux valeurs prédites par les calculs numériques et le modèle sur la Figure 5 . Il apparaît clairement que la gamme d’angles est la même. De plus, on observe une même tendance des valeurs des angles α mesurés dans les aquaporines à diminuer lorsque la longueur des cônes L augmente. Cette observation indique que les aquaporines possèdent une géométrie qui leur permet bien de minimiser la dissipation due aux effets d’entrée. Enfin, la comparaison quantitative de nos prédictions pour l’angle optimal et des angles mesurés dans les aquaporines semble indiquer la présence d’un glissement partiel au niveau des parois des cônes, avec des longueurs de glissement comprises entre 1 et 5a, soit entre 0,15 et 0,75 nm. | ||||||||
L’observation d’une protéine aux propriétés remarquables, l’aquaporine, nous a conduits à étudier l’influence de sa géométrie sur sa résistance hydrodynamique. Les profils réalisés par rayons X montrent en effet que ces petits canaux ont une forme proche de celle d’un sablier. Nos calculs numériques confirment que la géométrie de sablier joue bien un rôle important dans l’amélioration des performances de perméabilité à l’eau. La longueur des cônes, ainsi que la condition de glissement aux parois, influencent la valeur de l’angle optimum pour lequel la résistance hydrodynamique est minimale. Dans cette étude, nous appliquons l’hydrodynamique continue à l’aquaporine, dont les dimensions caractéristiques sont de l’ordre de la taille d’une molécule d’eau. Bien que l’on puisse s’attendre à un comportement différent du fait du caractère moléculaire de l’eau, il est apparu que notre approche restait pertinente. Pour nous assurer de cela, nous avons réalisé des simulations dites de dynamique moléculaire, pour cette fois étudier un écoulement de molécules discrètes à travers un bicône, et nous n’avons pas constaté de différence significatives avec la méthode continue [24]. Il est intéressant de constater que les aquaporines possèdent des paramètres géométriques qui lui confèrent le profil idéal pour réduire la dissipation, et donc maximiser le débit. Ce travail contribue ainsi à une meilleure compréhension des propriétés remarquables de ces protéines omniprésentes dans le corps humain. D’autres études devront permettre de comprendre l’influence du caractère moléculaire de l’eau sur l’écoulement, ainsi que l’influence de l’électrostatique ou encore de l’inhomogénéité de la surface des aquaporines. | ||||||||
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article. | ||||||||
Nous remercions A. Siria et A.-L. Biance pour d’intéressantes discussions. Cette recherche a été financée par le programme du European research council, projet Micromegas. | ||||||||
1.
Borgnia M, Nielsen S, Engel A, Agre P. Cellular and molecular biology of the aquaporin water channels . Annu Rev Biochem. 1999; ; 68 : :425.–458. 2.
Agre P. Aquaporin water channels (Nobel lecture) . Angew Chem Int Ed Engl. 2004; ; 43 : :4278.–4290. 3.
Bichet DG, Parent L, Sauvé R. Prix Nobel de Chimie 2003: canaux hydriques et ioniques . Med Sci (Paris). 2003; ; 19 : :1289.–1290. 4.
Guérin CF, Regli L, Badaut J. Rôles des aquaporines dans le cerveau . Med Sci (Paris). 2005; ; 21 : :747.–752. 5.
Murata K, Mitsuoka K, Hirai T, et al. Structural determinants of water permeation through aquaporin-1 . Nature. 2000; ; 407 : :599.–605. 6.
Sui H, Han BG, Lee JK, et al. Structural basis of water-specific transport through the AQP1 water channel . Nature. 2001; ; 414 : :872.–878. 7.
Bocquet L, Charlaix E. Nanofluidics, from bulk to interfaces . Chem Soc Rev. 2010; ; 39 : :1073.–1095. 8.
Majumder M, Chopra N, Andrews R, Hinds BJ. Nanoscale hydrodynamics . Enhanced flow in carbon nanotubes. Nature. 2005; ; 438 : :44.. 9.
Holt JK, Park HG, Wang YM, et al. Fast mass transport through sub-2-nanometer carbon nanotubes . Science. 2006; ; 312 : :1034.–1037. 10.
Cui Y, Bastien DA. Water transport in human aquaporin-4: molecular dynamics (MD) simulations . Biochem Biophys Res Commun. 2011; ; 412 : :654.–659. 11.
De Groot BL, Grubmüller H. Water permeation across biological membranes: mechanism and dynamics of aquaporin-1 and GlpF . Science. 2001; ; 294 : :2353.–2357. 12.
Gravelle S, Joly L, Detcheverry F, et al. Optimizing water permeability through the hourglass shape of aquaporins . Proc Natl Acad Sci USA. 2013; ; 110 : :2.–7. 13.
Bocquet L, J Barrat JL. Flow boundary conditions from nano- to micro-scales . Soft Matter. 2007; ; 3 : :685.–693. 14.
Sampson RA. On stokes’s current function . Philos Trans A Math Phys Eng Sci. 1891; ; 182 : :449.–518. 15.
Weissberg HL. End correction for slow viscous flow through long tubes . Phys Fluids. 1962; ; 5 : :1033.–1036. 17.
Hashido M, Kidera A, Ikeguchi M. Water transport in aquaporins: osmotic permeability matrix analysis of molecular dynamics simulations . Biophys J. 2007; ; 93 : :373.–385. 18.
Ho JD, Yeh R, Sandstrom A, et al. Crystal structure of human aquaporin 4 at 1.8 Å and its mechanism of conductance . Proc Natl Acad Sci USA. 2009;; 106 : :7437.–7442. 19.
De Groot BL, Engel A, Grubmüller H. The structure of the aquaporin-1 water channel: a comparison between cryo-electron microscopy and Xray crystallography . J Mol Biol. 2003; ; 325 : :485.–493. 20.
Harries WEC, Akhavan D, Miercke LJW, et al. The channel architecture of aquaporin 0 at a 2.2-Å resolution . Proc Natl Acad Sci USA. 2004;; 101 : :14045.–14050. 21.
Zhang YB, Chen LY. In silico study of aquaporin V: effects and affinity of the central pore-occluding lipid . Biophys Chem. 2013; ; 171 : :24.–30. 22.
Jensen MØ, Mouritsen OG. Single-channel water permeabilities of Escherichia coli aquaporins AqpZ and GlpF . Biophys J. 2006; ; 90 : :2270.–2284. | ||||||||