Théories explicatives de la dyslexie
2007
| ANALYSE |
18-
Apport de l’imagerie cérébrale
L'approche neurologique de la dyslexie repose sur les travaux fondateurs de Galaburda (1979 ) qui a découvert l'existence d'anomalies microscopiques dans la structure cyto-architectonique du cortex, en ayant l'opportunité d'analyser quelques cerveaux de personnes ayant présenté une probable dyslexie. Ce sont les progrès récents de la neuro-imagerie cérébrale qui ont permis l'accroissement des connaissances dans ce domaine et la confirmation de l'existence d'anomalies à la fois structurales et fonctionnelles du cerveau chez une majorité de personnes dyslexiques.
) qui a découvert l'existence d'anomalies microscopiques dans la structure cyto-architectonique du cortex, en ayant l'opportunité d'analyser quelques cerveaux de personnes ayant présenté une probable dyslexie. Ce sont les progrès récents de la neuro-imagerie cérébrale qui ont permis l'accroissement des connaissances dans ce domaine et la confirmation de l'existence d'anomalies à la fois structurales et fonctionnelles du cerveau chez une majorité de personnes dyslexiques.
) qui a découvert l'existence d'anomalies microscopiques dans la structure cyto-architectonique du cortex, en ayant l'opportunité d'analyser quelques cerveaux de personnes ayant présenté une probable dyslexie. Ce sont les progrès récents de la neuro-imagerie cérébrale qui ont permis l'accroissement des connaissances dans ce domaine et la confirmation de l'existence d'anomalies à la fois structurales et fonctionnelles du cerveau chez une majorité de personnes dyslexiques.Études anatomo-pathologiques ou morphologiques
Les études anatomo-pathologiques ou morphologiques réalisées chez des sujets adultes dyslexiques ont suggéré l'existence de particularités architecturales. Au niveau microscopique à partir de quelques cerveaux de sujets adultes dyslexiques, des anomalies (ectopies neuronales et dysplasies) retrouvées dans les régions péri-sylviennes gauches (Galaburda et coll., 1985) et dans le ganglion géniculé latéral (réduction de taille des neurones magnocellulaires) (Livingstone et coll., 1991) suggèrent l'existence d'un trouble de la migration et de la maturation neuronale. Ces résultats ont encouragé la recherche sur les bases neurologiques de la dyslexie. Les symptômes dyslexiques dans leur diversité pourraient dépendre de la localisation des anomalies micro-structurales, puisque ces dernières, variables dans leur localisation, pourraient venir perturber l'une ou l'autre des zones essentielles pour le transfert optimal de l'information à travers les réseaux sous-tendant les fonctions du langage ; chez l'enfant, cette perturbation pourrait concerner les réseaux de la lecture en voie de mise en place, nuisant à l'automatisation du transcodage des informations visuelles vers les régions codant pour la phonologie et réciproquement. À partir d'un modèle de souris, l'équipe de Galaburda (Jenner et coll., 2000) notait des anomalies de la connectivité secondaires à des ectopies neuronales. Des études morphométriques chez les dyslexiques ont rapporté des anomalies de l'architecture cérébrale (Habib, 2000 ; Leonard et coll., 2001 ; Rae et coll., 2002 ; Eckert et coll., 2003). Toutefois, la taille des effectifs, l'hétérogénéité phénotypique, la présence de comorbidités, ou les analyses a priori limitées à des régions d'intérêts rendent compte probablement des discordances retrouvées dans les résultats de ces études. Des méthodes récentes d'imagerie par résonance magnétique (IRM) ont permis certaines avancées. L'utilisation d'une méthode d'analyse statistique de la densité du signal IRM dans tout le volume cérébral (technique Voxel-Based Morphometry ou VBM) a suggéré selon plusieurs études des particularités du signal de la substance grise dans le cortex temporal, notamment dans l'hémisphère gauche ; les résultats de cette méthode restent toutefois d'interprétation délicate ; cette dernière est plus assurée lorsqu'elle peut s'appuyer sur d'autres sources d'informations, telles qu'une analyse focalisée dans une région montrant des anomalies fonctionnelles chez les mêmes sujets ou que l'établissement de corrélations entre données VBM et données comportementales (Silani et coll., 2005). Une autre méthode récente consiste dans une technique d'imagerie de diffusion : l'analyse de l'anisotropie du signal en imagerie de tenseur de diffusion. Par rapport à des sujets normolecteurs, une diminution du score d'anisotropie au niveau de fibres de la substance blanche sous-jacentes à la jonction temporo-pariétale gauche – zone essentielle pour la médiation phonologique de la lecture – a été montrée dans 3 études indépendantes chez des sujets d'âge variable, certains présentant des troubles de l'apprentissage de la lecture. Surtout, ces anomalies étaient souvent corrélées avec le score de lecture : une plus grande anisotropie, traduisant peut-être une moins bonne organisation anatomique au sein des faisceaux de fibres nerveuses concernés, tendant à être associée à de moins bonnes capacités de lecture (Klingberg et coll., 2000 ; Beaulieu et coll., 2005 ; Deutsch et coll., 2005). Il serait cependant important de montrer une corrélation de telles anomalies avec les scores de lecture au sein même de groupes de sujets dyslexiques, l'inclusion de sujets normolecteurs et de dyslexiques dans les études précédentes pouvant tendre à créer une distribution bimodale des performances et un effet de corrélation par construction.
) et dans le ganglion géniculé latéral (réduction de taille des neurones magnocellulaires) (Livingstone et coll., 1991) suggèrent l'existence d'un trouble de la migration et de la maturation neuronale. Ces résultats ont encouragé la recherche sur les bases neurologiques de la dyslexie. Les symptômes dyslexiques dans leur diversité pourraient dépendre de la localisation des anomalies micro-structurales, puisque ces dernières, variables dans leur localisation, pourraient venir perturber l'une ou l'autre des zones essentielles pour le transfert optimal de l'information à travers les réseaux sous-tendant les fonctions du langage ; chez l'enfant, cette perturbation pourrait concerner les réseaux de la lecture en voie de mise en place, nuisant à l'automatisation du transcodage des informations visuelles vers les régions codant pour la phonologie et réciproquement. À partir d'un modèle de souris, l'équipe de Galaburda (Jenner et coll., 2000) notait des anomalies de la connectivité secondaires à des ectopies neuronales. Des études morphométriques chez les dyslexiques ont rapporté des anomalies de l'architecture cérébrale (Habib, 2000 ; Leonard et coll., 2001 ; Rae et coll., 2002 ; Eckert et coll., 2003). Toutefois, la taille des effectifs, l'hétérogénéité phénotypique, la présence de comorbidités, ou les analyses a priori limitées à des régions d'intérêts rendent compte probablement des discordances retrouvées dans les résultats de ces études. Des méthodes récentes d'imagerie par résonance magnétique (IRM) ont permis certaines avancées. L'utilisation d'une méthode d'analyse statistique de la densité du signal IRM dans tout le volume cérébral (technique Voxel-Based Morphometry ou VBM) a suggéré selon plusieurs études des particularités du signal de la substance grise dans le cortex temporal, notamment dans l'hémisphère gauche ; les résultats de cette méthode restent toutefois d'interprétation délicate ; cette dernière est plus assurée lorsqu'elle peut s'appuyer sur d'autres sources d'informations, telles qu'une analyse focalisée dans une région montrant des anomalies fonctionnelles chez les mêmes sujets ou que l'établissement de corrélations entre données VBM et données comportementales (Silani et coll., 2005). Une autre méthode récente consiste dans une technique d'imagerie de diffusion : l'analyse de l'anisotropie du signal en imagerie de tenseur de diffusion. Par rapport à des sujets normolecteurs, une diminution du score d'anisotropie au niveau de fibres de la substance blanche sous-jacentes à la jonction temporo-pariétale gauche – zone essentielle pour la médiation phonologique de la lecture – a été montrée dans 3 études indépendantes chez des sujets d'âge variable, certains présentant des troubles de l'apprentissage de la lecture. Surtout, ces anomalies étaient souvent corrélées avec le score de lecture : une plus grande anisotropie, traduisant peut-être une moins bonne organisation anatomique au sein des faisceaux de fibres nerveuses concernés, tendant à être associée à de moins bonnes capacités de lecture (Klingberg et coll., 2000 ; Beaulieu et coll., 2005 ; Deutsch et coll., 2005). Il serait cependant important de montrer une corrélation de telles anomalies avec les scores de lecture au sein même de groupes de sujets dyslexiques, l'inclusion de sujets normolecteurs et de dyslexiques dans les études précédentes pouvant tendre à créer une distribution bimodale des performances et un effet de corrélation par construction.Imagerie fonctionnelle
L'imagerie fonctionnelle cérébrale permet de recueillir des indices de l'activité cérébrale dans l'ensemble du volume, avec une précision suffisante pour pouvoir reconstituer des images en coupes ou en volumes du cerveau et y localiser des régions dont on mesure le taux d'activité dans des conditions expérimentales diverses, incluant des tâches de lecture ou de perception phonologique du langage.
Considérations méthodologiques
L'évidence d'un dysfonctionnement des régions cérébrales impliquées dans la lecture et les processus cognitifs associés est venue des résultats des études en imagerie fonctionnelle. La tomographie d'émission de positons (TEP) ne peut être réalisée chez l'enfant dans le contexte d'examens à visée non diagnostique du fait de l'utilisation de radio-traceur même à faibles doses. Les méthodes de neuro-imagerie fonctionnelles sont un des principaux moyens d'exploration des corrélats neurobiologiques, mais les résultats de ces études nécessitent des interprétations prudentes par la possibilité de biais secondaires à des facteurs expérimentaux. Les corrélats cérébraux des fonctions linguistiques consistent en événements transitoires de faible amplitude et largement distribués dans tout le cerveau. Des variables telles que l'âge du sujet, la latéralité, la durée de présentation des stimuli et leur fréquence de présentation peuvent influencer les données recueillies et devront être contrôlées dans tout plan expérimental (Démonet et coll., 2005). Les études des troubles du langage en neuro-imagerie fonctionnelle sont particulièrement difficiles puisque vont intervenir des facteurs complexes induits par les troubles liés soit à l'anomalie cérébrale (exemple de l'aphasie), soit à la baisse des performances lors de la réalisation de la tâche. Dans le cas de la dyslexie, même s'il n'y a pas d'anomalie cérébrale, des facteurs spécifiques existent comme la diversité des déficits en termes de qualité et de sévérité et une sensibilité variable aux programmes de rééducation.
). Les études des troubles du langage en neuro-imagerie fonctionnelle sont particulièrement difficiles puisque vont intervenir des facteurs complexes induits par les troubles liés soit à l'anomalie cérébrale (exemple de l'aphasie), soit à la baisse des performances lors de la réalisation de la tâche. Dans le cas de la dyslexie, même s'il n'y a pas d'anomalie cérébrale, des facteurs spécifiques existent comme la diversité des déficits en termes de qualité et de sévérité et une sensibilité variable aux programmes de rééducation.Principaux résultats chez l'adulte et chez l'enfant
Dans les premières études avec les potentiels évoqués (PE), de simples tâches sensorielles furent principalement utilisées et des composants tardifs mesurés. Toutefois, dans les études plus récentes des tâches cognitives plus spécifiques guidées par les hypothèses théoriques ont permis de rapporter des déficits spécifiques aux sous-types de dyslexies (Taylor et Keenan, 1990 et 1999). Les enfants présentant un risque de développer une dyslexie peuvent montrer des anomalies des potentiels évoqués lors d'épreuves de lecture (Khan et coll., 1999), suggérant des processus corticaux différents dans ce trouble et soulignant la sensibilité de cette méthode dans l'investigation des troubles de la lecture (Taylor et Keenan, 1990 et 1999 ; Khan et coll., 1999 ; Breznitz et Leikin, 2000).
et 1999). Les enfants présentant un risque de développer une dyslexie peuvent montrer des anomalies des potentiels évoqués lors d'épreuves de lecture (Khan et coll., 1999), suggérant des processus corticaux différents dans ce trouble et soulignant la sensibilité de cette méthode dans l'investigation des troubles de la lecture (Taylor et Keenan, 1990 et 1999 ; Khan et coll., 1999 ; Breznitz et Leikin, 2000).Georgiewa et coll. (1999 et 2002) furent les premiers à rapporter des résultats, d'abord seulement en IRM fonctionnelle (IRMf) puis combinée à la méthode des PE chez des enfants dyslexiques et chez des témoins ; des différences entre les deux groupes étaient retrouvées pour des tâches linguistiques avec une présentation visuelle des stimuli au niveau du cortex frontal inférieur gauche, entre 250 et 600 ms (Georgiewa et coll., 2002). À partir d'une tâche de détection de trait ou de mot, Helenius et coll. (1999) montraient qu'il existe chez les sujets dyslexiques un déficit dans le traitement pré-lexical impliquant habituellement le cortex temporo-occipital inférieur gauche. La présence d'une « N100m » (événement magnétique évoqué de latence 100 ms) normale était en faveur de la normalité des processus précoces du traitement visuel. Cette dernière caractéristique est cependant incertaine, puisque la même équipe a au contraire rapporté des anomalies des composantes précoces, mais dans le domaine de la perception auditive (Helenius et coll., 2002).
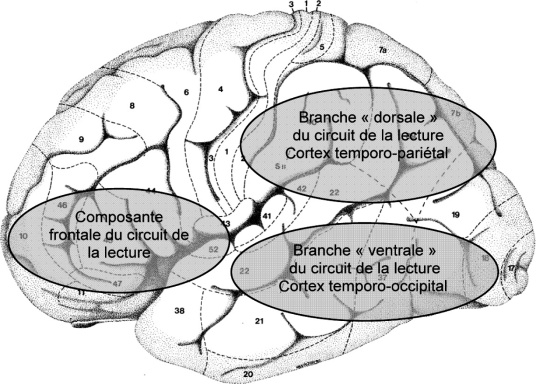
et 2002) furent les premiers à rapporter des résultats, d'abord seulement en IRM fonctionnelle (IRMf) puis combinée à la méthode des PE chez des enfants dyslexiques et chez des témoins ; des différences entre les deux groupes étaient retrouvées pour des tâches linguistiques avec une présentation visuelle des stimuli au niveau du cortex frontal inférieur gauche, entre 250 et 600 ms (Georgiewa et coll., 2002). À partir d'une tâche de détection de trait ou de mot, Helenius et coll. (1999) montraient qu'il existe chez les sujets dyslexiques un déficit dans le traitement pré-lexical impliquant habituellement le cortex temporo-occipital inférieur gauche. La présence d'une « N100m » (événement magnétique évoqué de latence 100 ms) normale était en faveur de la normalité des processus précoces du traitement visuel. Cette dernière caractéristique est cependant incertaine, puisque la même équipe a au contraire rapporté des anomalies des composantes précoces, mais dans le domaine de la perception auditive (Helenius et coll., 2002).Chez l'adulte normolecteur, les régions cérébrales impliquées dans la lecture de mots isolés sont largement distribuées mais dominées par un réseau hémisphérique gauche avec deux circuits postérieurs et un circuit antérieur (Pugh et coll., 2000). Le circuit ventral ou temporo-occipital est centré sur le gyrus fusiforme (Visual Word Form Area) et semble sous-tendre la procédure d'adressage ou d'accès quasi automatique à la forme visuelle des mots (Cohen et coll., 2002) ; le circuit dorsal ou temporo-pariétal comprend principalement le gyrus angulaire et le gyrus supra-marginalis (Price, 1998) impliqué dans le traitement phonologique et le processus, plus lent, d'assemblage. Le circuit antérieur est quant à lui centré principalement sur le gyrus frontal inférieur gauche ; il est relié aux deux circuits postérieurs (Price et coll., 2001) et est impliqué dans les processus phonologique et articulatoire lors de la phase de production des mots.
). Le circuit ventral ou temporo-occipital est centré sur le gyrus fusiforme (Visual Word Form Area) et semble sous-tendre la procédure d'adressage ou d'accès quasi automatique à la forme visuelle des mots (Cohen et coll., 2002) ; le circuit dorsal ou temporo-pariétal comprend principalement le gyrus angulaire et le gyrus supra-marginalis (Price, 1998) impliqué dans le traitement phonologique et le processus, plus lent, d'assemblage. Le circuit antérieur est quant à lui centré principalement sur le gyrus frontal inférieur gauche ; il est relié aux deux circuits postérieurs (Price et coll., 2001) et est impliqué dans les processus phonologique et articulatoire lors de la phase de production des mots.Chez les adultes dyslexiques comparativement à des sujets témoins normo-lecteurs est mise en évidence une réduction de l'activité des circuits postérieurs (figure 18.1). La région clé du circuit dorsal, le gyrus angulaire, montre une corrélation positive de son niveau d'activation avec les scores de lecture chez les sujets normolecteurs et une corrélation négative chez les dyslexiques (Rumsey et coll., 1999). Une étude en TEP (Paulesu et coll., 2001) chez des adultes dyslexiques bien compensés et des témoins, de trois nationalités différentes (anglaise, italienne et française) retrouvait un défaut d'activation chez les dyslexiques du circuit ventral quelque soit la langue. Ces résultats rejoignent ceux rapportés dans des études réalisées avec la magnéto-encéphalographie (Helenius et coll., 1999). Bien que la région frontale inférieure gauche et des régions hémisphériques droites semblent moins actives chez les dyslexiques dans certaines études (Paulesu et coll., 1996 ; Rumsey et coll., 1997), dans d'autres au contraire les auteurs retrouvaient une augmentation d'activité suggérant des mécanismes de compensation du cortex prémoteur visant à suppléer le dysfonctionnement des régions postérieures gauches : au niveau du cortex frontal gauche (Shaywitz et coll., 1998 ; Brunswick et coll., 1999 ; Georgiewa et coll., 2002) et au niveau des régions droites (Simos et coll., 2002a).
). La région clé du circuit dorsal, le gyrus angulaire, montre une corrélation positive de son niveau d'activation avec les scores de lecture chez les sujets normolecteurs et une corrélation négative chez les dyslexiques (Rumsey et coll., 1999). Une étude en TEP (Paulesu et coll., 2001) chez des adultes dyslexiques bien compensés et des témoins, de trois nationalités différentes (anglaise, italienne et française) retrouvait un défaut d'activation chez les dyslexiques du circuit ventral quelque soit la langue. Ces résultats rejoignent ceux rapportés dans des études réalisées avec la magnéto-encéphalographie (Helenius et coll., 1999). Bien que la région frontale inférieure gauche et des régions hémisphériques droites semblent moins actives chez les dyslexiques dans certaines études (Paulesu et coll., 1996 ; Rumsey et coll., 1997), dans d'autres au contraire les auteurs retrouvaient une augmentation d'activité suggérant des mécanismes de compensation du cortex prémoteur visant à suppléer le dysfonctionnement des régions postérieures gauches : au niveau du cortex frontal gauche (Shaywitz et coll., 1998 ; Brunswick et coll., 1999 ; Georgiewa et coll., 2002) et au niveau des régions droites (Simos et coll., 2002a).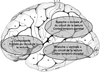
Les études développementales en imagerie fonctionnelle (tableau 18.I) permettent d'interpréter et comprendre certains résultats discordants des études chez l'adulte. Shaywitz et coll. (2002) étudiant 144 enfants dyslexiques et témoins montraient que l'activité augmente avec l'âge dans les régions frontales inférieures gauches et droites chez les sujets dyslexiques lors d'une tâche de rimes. Ces résultats sont en faveur de l'hypothèse compensatoire : l'augmentation de l'activation dans les régions frontales et/ou les régions hémisphériques droites constitue un moyen de surmonter le défaut d'engagement des régions postérieures gauches. Ces chercheurs (Shaywitz et coll., 2002) suggéraient également que la diminution de l'activité cérébrale dans le circuit ventral chez l'adulte dyslexique n'est pas la conséquence des difficultés persistantes pour la lecture puisqu'ils retrouvaient une corrélation positive entre l'activité de cette aire et le niveau de lecture chez l'enfant dyslexique. Toutefois, Simos et coll. (2000a et b) rapportaient que cette région chez les enfants dyslexiques présente un niveau d'activité comparable aux témoins et que la différence se situe essentiellement au niveau du décours temporel de l'engagement de ces aires cérébrales : l'activité neuronale 250 à 1 200 ms après présentation du stimulus « saute » vers le cortex temporal droit chez l'enfant dyslexique (Simos et coll., 2000c) alors que chez les normolecteurs elle se propage vers les régions temporale et pariétale gauches. Au total, ces résultats suggèrent une anomalie de la connectivité au sein des circuits temporo-pariéto-frontaux qui sous-tendent le langage et affectant peut-être particulièrement le circuit sous-jacent à la boucle phonologique en mémoire de travail ainsi que l'interaction entre les « circuits dorsal et ventral de la lecture » (selon la conceptualisation de Pugh et coll., 2000), ce qui rejoint les conclusions d'autres études comme celle d'Horwitz et coll. (1998) ou de Klingberg et coll. (2000).
) permettent d'interpréter et comprendre certains résultats discordants des études chez l'adulte. Shaywitz et coll. (2002) étudiant 144 enfants dyslexiques et témoins montraient que l'activité augmente avec l'âge dans les régions frontales inférieures gauches et droites chez les sujets dyslexiques lors d'une tâche de rimes. Ces résultats sont en faveur de l'hypothèse compensatoire : l'augmentation de l'activation dans les régions frontales et/ou les régions hémisphériques droites constitue un moyen de surmonter le défaut d'engagement des régions postérieures gauches. Ces chercheurs (Shaywitz et coll., 2002) suggéraient également que la diminution de l'activité cérébrale dans le circuit ventral chez l'adulte dyslexique n'est pas la conséquence des difficultés persistantes pour la lecture puisqu'ils retrouvaient une corrélation positive entre l'activité de cette aire et le niveau de lecture chez l'enfant dyslexique. Toutefois, Simos et coll. (2000a et b) rapportaient que cette région chez les enfants dyslexiques présente un niveau d'activité comparable aux témoins et que la différence se situe essentiellement au niveau du décours temporel de l'engagement de ces aires cérébrales : l'activité neuronale 250 à 1 200 ms après présentation du stimulus « saute » vers le cortex temporal droit chez l'enfant dyslexique (Simos et coll., 2000c) alors que chez les normolecteurs elle se propage vers les régions temporale et pariétale gauches. Au total, ces résultats suggèrent une anomalie de la connectivité au sein des circuits temporo-pariéto-frontaux qui sous-tendent le langage et affectant peut-être particulièrement le circuit sous-jacent à la boucle phonologique en mémoire de travail ainsi que l'interaction entre les « circuits dorsal et ventral de la lecture » (selon la conceptualisation de Pugh et coll., 2000), ce qui rejoint les conclusions d'autres études comme celle d'Horwitz et coll. (1998) ou de Klingberg et coll. (2000).Tableau 18.I Études en imagerie fonctionnelle chez les enfants dyslexiques (Dys) comparés aux témoins normolecteurs (NL)
|
Études
|
Technique d'imagerie
|
Population (âge en années)
|
Tâches cognitives
|
Principaux résultats chez les Dys comparés aux NL
|
|---|---|---|---|---|
|
Georgiewa et coll., 1999
|
IRMf
|
17 Dys (9-17)
17 NL (9-17) |
Paradigme visuel hiérarchique : processus orthographique, phonologique et sémantique
|
Diminution de l'activation des régions frontale et temporale inférieure gauches lors de la lecture de pseudo-mots et d'une tâche métaphonologique
|
|
Richards et coll., 1999
|
sIRM
|
6 Dys (9-12)
7 NL (9-12) |
Tâche auditive de rimes
|
Augmentation du métabolisme du lactate dans les régions frontales gauches
|
|
Simos et coll., 2000a
|
MEG
|
10 Dys (10-17)
8 NL (8-16) |
Tâches visuelle et auditive de reconnaissance de mots
|
Activation normale de la région temporale inférieure gauche
Diminution de l'activité de la région temporo-pariétale gauche et augmentation dans la région homologue droite lors de la lecture de mots |
|
Simos et coll., 2000b
|
MEG
|
11 Dys (10-17)
10 NL (8-16) |
Tâche visuelle de rimes avec des pseudo-mots
|
Activation normale de la région temporale inférieure gauche
Diminution de l'activité de la région temporo-pariétale gauche et augmentation dans la région homologue droite lors de la lecture de pseudo-mots |
|
Corina et coll., 2001
|
IRMf
|
8 Dys (10-13)
8 NL (10-13) |
Tâche auditive de jugement phonologique et lexical
|
Augmentation supérieure de l'activité de la région du gyrus temporal inférieur droit par rapport aux gyrus temporal inférieur et précentral gauches, lors d'une tâche de jugement phonologique
Diminution de l'activité du gyrus frontal moyen de façon bilatérale et augmentation de l'activité au niveau du cortex fronto-orbitaire gauche lors d'une tâche de jugement lexical |
|
Temple et coll., 2001
|
IRMf
|
24 Dys (8-12)
15 NL (8-12) |
Tâches visuelles : traitement phonologique et orthographique
|
Activation normale de la région frontale gauche et diminuée dans la région temporo-pariétale gauche lors d'une tâche de jugement de rimes
|
|
Shaywitz et coll., 2002
|
IRMf
|
70 Dys (7-18)
74 NL (7-17) |
Paradigme visuel hiérarchique : processus orthographique, phonologique et sémantique
|
Réduction d'activité dans les régions pariéto-temporale et occipito-temporale gauches lors de processus impliqués dans la lecture
Corrélation entre l'âge et le niveau d'activation des régions frontales droite et gauche (augmentation avec l'âge des sujets) |
|
Georgiewa et coll., 2002
|
IRMf PE
|
9 Dys (12,6)
8 NL (12,7) |
Lecture silencieuse de mots et de non-mots
|
Augmentation de l'activation au niveau du gyrus frontal gauche
|
|
IRMf : IRM focntionelle ; sIRM : IRM spectroscopique ; MEG : magneto-encéphalographique ; PE : potentiels evoqués
|
||||
Imagerie fonctionnelle et test des hypothèses physiopathologiques des dyslexies
Les nombreuses hypothèses physiopathologiques concernant l'origine des dyslexies ont toutes reçu un support empirique d'études de neuro-imagerie confirmant l'existence d'effets différentiels prédits. Ces hypothèses peuvent être classifiées en deux types. Le premier type suppose l'existence d'un déficit affectant les capacités de traitement explicite de représentations phonologiques ou lexicales stockées en mémoire à long terme. Le second type recouvre de nombreuses hypothèses supposant des anomalies diverses affectant des processus implicites, de type sensorimoteur, permettant le traitement en temps réel de l'information perceptive : déficit dans le système magnocellulaire, déficit d'orientation automatique de l'attention, déficit de l'adaptation sensori-motrice au niveau cérébelleux.
Dans la mesure où le déficit phonologique constitue un facteur étiologique prédominant dans la dyslexie, la majorité des études en imagerie fonctionnelle ont évalué ces processus à partir de tâches de rimes (Rumsey et coll., 1997 ; Simos et coll., 2000b), de tâches de mémoire verbale de travail (Paulesu et coll., 1996) ou lors de la présentation auditive de stimuli verbaux (Rumsey et coll., 1992 ; Simos et coll., 2000a). La plupart des études ont montré un hypofonctionnement des régions périsylviennes plutôt gauches. Paulesu et coll. (1996) suggéraient que le pattern retrouvé chez les sujets dyslexiques peut être en rapport avec une déconnexion dans le circuit péri-sylvien gauche. Une équipe a étudié en IRMf la perception catégorielle des phonèmes. Les sujets dyslexiques présentaient une diminution de l'activité dans le gyrus supra-marginalis gauche (Ruff et coll., 2002), une région clé dans les processus phonologiques (Démonet et coll., 1996).
; Simos et coll., 2000b), de tâches de mémoire verbale de travail (Paulesu et coll., 1996) ou lors de la présentation auditive de stimuli verbaux (Rumsey et coll., 1992 ; Simos et coll., 2000a). La plupart des études ont montré un hypofonctionnement des régions périsylviennes plutôt gauches. Paulesu et coll. (1996) suggéraient que le pattern retrouvé chez les sujets dyslexiques peut être en rapport avec une déconnexion dans le circuit péri-sylvien gauche. Une équipe a étudié en IRMf la perception catégorielle des phonèmes. Les sujets dyslexiques présentaient une diminution de l'activité dans le gyrus supra-marginalis gauche (Ruff et coll., 2002), une région clé dans les processus phonologiques (Démonet et coll., 1996).Dans plusieurs études, des arguments concordants pour un dysfonctionnement du système visuel magnocellulaire ont été rapportés. Par exemple, Eden et coll. (1996) retrouvaient un défaut d'activation de l'aire MT/V5 durant une tâche de détection de mouvement et mettaient en évidence une corrélation entre le niveau d'activation de l'aire V5 et la vitesse de lecture chez les dyslexiques et les sujets normo-lecteurs. Toutefois, d'autres études n'ont pas confirmé ces résultats (Johannes et coll., 1996 ; Vanni et coll., 1997 ; Amitay et coll., 2002). Les effets du système magnocellulaire paraissent en fait subtils et mis en évidence uniquement sous certaines contraintes expérimentales (Bednareck et Grabowska, 2002).
) retrouvaient un défaut d'activation de l'aire MT/V5 durant une tâche de détection de mouvement et mettaient en évidence une corrélation entre le niveau d'activation de l'aire V5 et la vitesse de lecture chez les dyslexiques et les sujets normo-lecteurs. Toutefois, d'autres études n'ont pas confirmé ces résultats (Johannes et coll., 1996 ; Vanni et coll., 1997 ; Amitay et coll., 2002). Les effets du système magnocellulaire paraissent en fait subtils et mis en évidence uniquement sous certaines contraintes expérimentales (Bednareck et Grabowska, 2002).Temple et coll. (2000) étudiaient en IRMf la réponse cérébrale à des variations rapides de la trace acoustique du signal de parole et montraient chez les sujets normo-lecteurs une augmentation de l'activation au niveau de la région frontale inférieure gauche et de l'hémisphère cérébelleux droit lors de variations rapides comparativement à des changements plus lents du signal acoustique. Chez les sujets dyslexiques, les auteurs notaient une augmentation de l'activation dans ces mêmes régions pour des variations lentes. De façon similaire, Nagarajan et coll. (1999) retrouvaient une diminution de la M100 obtenue en MEG en réponse à des stimuli acoustiques présentant des changements rapides chez des sujets dyslexiques alors qu'ils présentaient une M100 d'amplitude plus large que les témoins pour des stimuli plus lents.
) retrouvaient une diminution de la M100 obtenue en MEG en réponse à des stimuli acoustiques présentant des changements rapides chez des sujets dyslexiques alors qu'ils présentaient une M100 d'amplitude plus large que les témoins pour des stimuli plus lents.Ces effets étaient également étudiés avec des syllabes naturelles (ma/na), qu'elles soient modifiées ou non par un étirement des transitions formantiques (Ruff et coll., 2002). Les régions cérébrales sensibles aux changements acoustiques étaient identifiées comme étant la région frontale gauche et le ralentissement de la parole entraînait une augmentation d'activation dans cette région pour les sujets dyslexiques. Mais le niveau d'activation de la région du gyrus supra-marginalis gauche n'était pas influencé par ces variations de signal acoustique, alors que cette région est habituellement le siège d'un défaut d'activation chez les adultes dyslexiques. Deux effets importants impliqués dans les mécanismes de base de la dyslexie étaient identifiés par cette étude : l'activité neuronale était augmentée par le ralentissement de la parole dans certaines régions cérébrales et le défaut d'activité neuronale dans l'aire du gyrus supra-marginalis était le support du déficit phonologique dans la dyslexie. Les variations topographiques de l'activation cérébrale dans cette étude pourraient rendre compte de certains résultats discordants voire contradictoires retrouvés dans diverses expérimentations et constituent un exemple dans lequel les résultats d'une étude peuvent vérifier et/ou compléter des résultats obtenus par d'autres équipes.
). Les régions cérébrales sensibles aux changements acoustiques étaient identifiées comme étant la région frontale gauche et le ralentissement de la parole entraînait une augmentation d'activation dans cette région pour les sujets dyslexiques. Mais le niveau d'activation de la région du gyrus supra-marginalis gauche n'était pas influencé par ces variations de signal acoustique, alors que cette région est habituellement le siège d'un défaut d'activation chez les adultes dyslexiques. Deux effets importants impliqués dans les mécanismes de base de la dyslexie étaient identifiés par cette étude : l'activité neuronale était augmentée par le ralentissement de la parole dans certaines régions cérébrales et le défaut d'activité neuronale dans l'aire du gyrus supra-marginalis était le support du déficit phonologique dans la dyslexie. Les variations topographiques de l'activation cérébrale dans cette étude pourraient rendre compte de certains résultats discordants voire contradictoires retrouvés dans diverses expérimentations et constituent un exemple dans lequel les résultats d'une étude peuvent vérifier et/ou compléter des résultats obtenus par d'autres équipes.La MMN (Mitch Match Negativity) joue un rôle important dans l'exploration de l'hypothèse de déficits perceptifs. Cette composante neurophysiologique est obtenue lors de tout changement identifiable lors d'une tâche auditive de répétition de stimuli indépendamment du niveau d'attention du sujet (Kujala et Naatanen, 2001), suggérant un processus de type automatique. Les études initiales chez l'enfant présentant un trouble des apprentissages montraient une diminution de l'amplitude de la MMN mais la spécificité de cet effet paraît incertaine : la diminution de la MMN est liée au caractère linguistique des stimuli pour Schulte-Körne et coll. (1998), l'anomalie de la MMN semble sensible à un facteur phonétique particulier, le temps de voisement (différence dans le décours temporel des événements acoustiques successifs entre « ga » et « ka » par exemple) pour Kraus et coll. (1996), alors que Baldeweg et coll. (1999) identifie cette anomalie lors d'une condition faisant varier un facteur non pas temporel mais spectral : la hauteur tonale. Ces résultats pourraient refléter des corrélats neurophysiologiques des troubles de la discrimination des sons de la parole chez le dyslexique, bien que le déterminisme de ces troubles paraisse donc variable. Il faut noter que Giraud et coll. (2005) ont confirmé l'existence d'un trouble du traitement temporel du signal de parole (absence d'une composante électrophysiologique caractéristique en français du son de pré-voisement du phonème « b ») mais seulement dans un sous-groupe de dyslexiques présentant des troubles sévères, et alors qu'un autre sous-groupe également sévèrement déficitaire présentait un profil électrophysiologique caractérisé non par l'absence de cette composante mais par des composantes multiples. Une partie de cette même étude, non encore disponible sous forme de publication, montre en outre un profil électrophysiologique normal dans un 3e sous-groupe, composé de patients moins sévèrement déficitaires. Des paradigmes similaires ont permis à Leppanen et coll. (1999) de comparer des enfants à risque et non à risque de développer une dyslexie en fonction de leur histoire familiale : les résultats de ces études suggèrent que dès l'âge de 6 mois les sujets avec des antécédents familiaux de dyslexie traitent les stimuli différemment.
), suggérant un processus de type automatique. Les études initiales chez l'enfant présentant un trouble des apprentissages montraient une diminution de l'amplitude de la MMN mais la spécificité de cet effet paraît incertaine : la diminution de la MMN est liée au caractère linguistique des stimuli pour Schulte-Körne et coll. (1998), l'anomalie de la MMN semble sensible à un facteur phonétique particulier, le temps de voisement (différence dans le décours temporel des événements acoustiques successifs entre « ga » et « ka » par exemple) pour Kraus et coll. (1996), alors que Baldeweg et coll. (1999) identifie cette anomalie lors d'une condition faisant varier un facteur non pas temporel mais spectral : la hauteur tonale. Ces résultats pourraient refléter des corrélats neurophysiologiques des troubles de la discrimination des sons de la parole chez le dyslexique, bien que le déterminisme de ces troubles paraisse donc variable. Il faut noter que Giraud et coll. (2005) ont confirmé l'existence d'un trouble du traitement temporel du signal de parole (absence d'une composante électrophysiologique caractéristique en français du son de pré-voisement du phonème « b ») mais seulement dans un sous-groupe de dyslexiques présentant des troubles sévères, et alors qu'un autre sous-groupe également sévèrement déficitaire présentait un profil électrophysiologique caractérisé non par l'absence de cette composante mais par des composantes multiples. Une partie de cette même étude, non encore disponible sous forme de publication, montre en outre un profil électrophysiologique normal dans un 3e sous-groupe, composé de patients moins sévèrement déficitaires. Des paradigmes similaires ont permis à Leppanen et coll. (1999) de comparer des enfants à risque et non à risque de développer une dyslexie en fonction de leur histoire familiale : les résultats de ces études suggèrent que dès l'âge de 6 mois les sujets avec des antécédents familiaux de dyslexie traitent les stimuli différemment.Nicolson et coll. (1999) ont utilisé la TEP lors d'un paradigme d'apprentissage d'une séquence de mouvements des doigts pour évaluer l'activation cérébrale chez des sujets dyslexiques et témoins. Ils montraient une diminution de l'activation cérébrale au niveau de l'hémisphère cérébelleux droit et du cortex frontal inférieur gauche chez les sujets dyslexiques par rapport aux témoins. Ces anomalies corroborent les anomalies métaboliques au niveau de l'hémisphère cérébelleux droit retrouvées chez les dyslexiques lors d'une étude en spectro-IRM (Rae et coll., 1998). Néanmoins, si des anomalies de l'activation des régions cérébelleuses sont parfois retrouvées lors d'études en imagerie fonctionnelle chez les sujets dyslexiques, elles ne sont pas toujours interprétées (Brunswick et coll., 1999).
) ont utilisé la TEP lors d'un paradigme d'apprentissage d'une séquence de mouvements des doigts pour évaluer l'activation cérébrale chez des sujets dyslexiques et témoins. Ils montraient une diminution de l'activation cérébrale au niveau de l'hémisphère cérébelleux droit et du cortex frontal inférieur gauche chez les sujets dyslexiques par rapport aux témoins. Ces anomalies corroborent les anomalies métaboliques au niveau de l'hémisphère cérébelleux droit retrouvées chez les dyslexiques lors d'une étude en spectro-IRM (Rae et coll., 1998). Néanmoins, si des anomalies de l'activation des régions cérébelleuses sont parfois retrouvées lors d'études en imagerie fonctionnelle chez les sujets dyslexiques, elles ne sont pas toujours interprétées (Brunswick et coll., 1999).Globalement, l'ensemble très important de données accumulées ces dernières années est caractérisé par la grande diversité des anomalies rapportées dans la mesure où des publications sont venues à l'appui de chacune des théories concurrentes pour rendre compte de la physiopathologie des dyslexies. Cette diversité des effets décrits contraste avec la relative constance des résultats concernant les expériences d'imagerie ayant utilisé des tâches de lecture. Elle plaide aussi en faveur de l'hétérogénéité phénotypique des dyslexies.
Rééducations et bases neurologiques
Associé à une méthode d'enseignement appropriée, le principe des rééducations repose sur des interventions sur le langage, la phonologie, la lecture ou la parole adaptées au handicap de l'enfant. Bien qu'un certain degré de compensation puisse survenir avec le temps, la dyslexie est généralement un trouble persistant (Shaywitz et coll., 1999) qui peut être à l'origine d'un retentissement sévère sur la progression académique de l'enfant. Les traitements comportent souvent des entraînements phonologiques mais peuvent également inclure d'autres dimensions des processus linguistiques comme la morphologie, la syntaxe ou le niveau discursif. Peu de méthodes d'entraînement commercialisées et utilisées en pratique clinique ont été évaluées pour leur pertinence. Malgré différentes modalités et durée de remédiation, les études concernant les méthodes basées sur les aspects phonologiques ont montré une amélioration des capacités phonologiques après un entraînement intensif (Hatcher et Hume, 1999 ; Torgensen et coll., 1999 ; Wise et coll., 2000). Néanmoins, la généralisation des effets obtenus aux capacités de lecture est inconstante ; le succès d'une méthode est sous la dépendance de différences individuelles et les facteurs prédictifs d'une bonne réponse à la remédiation restent à déterminer.
) qui peut être à l'origine d'un retentissement sévère sur la progression académique de l'enfant. Les traitements comportent souvent des entraînements phonologiques mais peuvent également inclure d'autres dimensions des processus linguistiques comme la morphologie, la syntaxe ou le niveau discursif. Peu de méthodes d'entraînement commercialisées et utilisées en pratique clinique ont été évaluées pour leur pertinence. Malgré différentes modalités et durée de remédiation, les études concernant les méthodes basées sur les aspects phonologiques ont montré une amélioration des capacités phonologiques après un entraînement intensif (Hatcher et Hume, 1999 ; Torgensen et coll., 1999 ; Wise et coll., 2000). Néanmoins, la généralisation des effets obtenus aux capacités de lecture est inconstante ; le succès d'une méthode est sous la dépendance de différences individuelles et les facteurs prédictifs d'une bonne réponse à la remédiation restent à déterminer.L'imagerie fonctionnelle (IRMf et MEG1
) permet non seulement d'étudier les déficits des réseaux neuronaux impliqués dans les fonctions linguistiques mais est aussi un moyen d'évaluation de la plasticité cérébrale qui fait suite à des phénomènes de compensation (Shaywitz et coll., 2003) ou à des entraînements intensifs que ce soit chez l'adulte (Eden et coll., 2004) ou chez l'enfant. Dans six études réalisées chez des enfants dyslexiques (tableau 18.II), une corrélation entre le fonctionnement cérébral et les améliorations des performances linguistiques était démontrée avec des protocoles de rééducation divers mais cependant tous caractérisés par le caractère intensif de la prise en charge, sur des périodes variables de temps. La plus grande influence sur les performances et les signaux neurofonctionnels d'une intervention phonologique intensive par rapport à une prise en charge conventionnelle a été montrée dans certaines de ces études (Shaywitz et coll., 2004). Dans l'étude réalisée par Temple et coll. (2003), les enfants dyslexiques n'activaient pas, avant entraînement, leur région temporo-pariétale gauche, région activée chez les normo-lecteurs par une tâche de jugement de rimes. En revanche, les deux groupes activaient la région frontale gauche mais dans des aires différentes. Après entraînement, les sujets dyslexiques montraient une augmentation de l'activité dans la région temporale ou pariétale gauche sans que cette activation recouvre la région initialement hypofonctionnelle et une activation dans la région frontale gauche dans la même aire que les témoins. Des augmentations d'activité étaient également montrées dans des régions temporale et frontale droites. Des corrélations positives étaient retrouvées entre les modifications du signal IRM dans les régions temporo-pariétale gauche et frontale droite et les changements pour les scores de langage ou des aptitudes phonologiques. Toutefois, il n'était pas mis en évidence de corrélation avec les scores de lecture. Les différentes techniques d'imagerie fonctionnelle sont certes suffisamment sensibles pour mettre en évidence des modifications de l'activité cérébrale après des remédiations, mais la spécificité des mécanismes à l'origine des effets obtenus tant au niveau cognitif que cérébral doit être discutée. Bien qu'il semble aisé d'obtenir des arguments en faveur d'une certaine plasticité cérébrale après entraînement, le fait d'obtenir les mêmes améliorations après des entraînements aussi bien phonologique (Simos et coll., 2002a) que non verbal (Kujala et coll., 2001) laisse perplexe quant au caractère spécifique de cet effet. Au cours des entraînements, plusieurs facteurs, spécifiques et non spécifiques du langage, semblent avoir une influence sur les symptômes de la dyslexie et leur base neurologique. Comment l'imagerie fonctionnelle peut-elle nous aider à comprendre la dynamique et les mécanismes des changements induits par les remédiations ? Kujala et coll. (2001) n'établissaient pas de lien direct entre l'entraînement audio-visuel et un test de lecture, alors que d'autres chercheurs ont mis à profit le lien entre le déficit phonologique testé, la méthode de rééducation et le paradigme d'imagerie (Simos et coll., 2002a ; Temple et coll., 2003).
) ou à des entraînements intensifs que ce soit chez l'adulte (Eden et coll., 2004) ou chez l'enfant. Dans six études réalisées chez des enfants dyslexiques (tableau 18.II), une corrélation entre le fonctionnement cérébral et les améliorations des performances linguistiques était démontrée avec des protocoles de rééducation divers mais cependant tous caractérisés par le caractère intensif de la prise en charge, sur des périodes variables de temps. La plus grande influence sur les performances et les signaux neurofonctionnels d'une intervention phonologique intensive par rapport à une prise en charge conventionnelle a été montrée dans certaines de ces études (Shaywitz et coll., 2004). Dans l'étude réalisée par Temple et coll. (2003), les enfants dyslexiques n'activaient pas, avant entraînement, leur région temporo-pariétale gauche, région activée chez les normo-lecteurs par une tâche de jugement de rimes. En revanche, les deux groupes activaient la région frontale gauche mais dans des aires différentes. Après entraînement, les sujets dyslexiques montraient une augmentation de l'activité dans la région temporale ou pariétale gauche sans que cette activation recouvre la région initialement hypofonctionnelle et une activation dans la région frontale gauche dans la même aire que les témoins. Des augmentations d'activité étaient également montrées dans des régions temporale et frontale droites. Des corrélations positives étaient retrouvées entre les modifications du signal IRM dans les régions temporo-pariétale gauche et frontale droite et les changements pour les scores de langage ou des aptitudes phonologiques. Toutefois, il n'était pas mis en évidence de corrélation avec les scores de lecture. Les différentes techniques d'imagerie fonctionnelle sont certes suffisamment sensibles pour mettre en évidence des modifications de l'activité cérébrale après des remédiations, mais la spécificité des mécanismes à l'origine des effets obtenus tant au niveau cognitif que cérébral doit être discutée. Bien qu'il semble aisé d'obtenir des arguments en faveur d'une certaine plasticité cérébrale après entraînement, le fait d'obtenir les mêmes améliorations après des entraînements aussi bien phonologique (Simos et coll., 2002a) que non verbal (Kujala et coll., 2001) laisse perplexe quant au caractère spécifique de cet effet. Au cours des entraînements, plusieurs facteurs, spécifiques et non spécifiques du langage, semblent avoir une influence sur les symptômes de la dyslexie et leur base neurologique. Comment l'imagerie fonctionnelle peut-elle nous aider à comprendre la dynamique et les mécanismes des changements induits par les remédiations ? Kujala et coll. (2001) n'établissaient pas de lien direct entre l'entraînement audio-visuel et un test de lecture, alors que d'autres chercheurs ont mis à profit le lien entre le déficit phonologique testé, la méthode de rééducation et le paradigme d'imagerie (Simos et coll., 2002a ; Temple et coll., 2003).Tableau 18.II Effets des méthodes d'entraînement intensif chez les enfants dyslexiques (Dys) comparés à des sujets témoins (NL)
|
Études
|
Technique Imagerie
|
Population (âge en années)
|
Méthode d'entraînement
|
Principaux résultats après entraînement
|
|---|---|---|---|---|
|
Richards et coll., 2000
|
sIRMf
|
8 Dys (10,6)
7 NL (10,3) |
Entraînement phonologique
3 semaines 15 sessions de 2 heures |
Amélioration des performances phonologiques
Diminution du métabolisme du lactate dans la région antérieure gauche |
|
Kujala et coll., 2001
|
PEs (MMN)
|
24 Dys (7)
24 Dys (7) 11 Dys (7) 11 Dys témoins (7) |
Entraînement audio-visuel non verbal
7 semaines 2 sessions de 10 min par semaine |
Amélioration des capacités de lecture
Augmentation de l'amplitude de la MMN Corrélation avec les modifications des performances |
|
Simos et coll., 2002a
|
MEG
|
8 Dys (11,4)
8 NL (10,3) |
Entraînement phonologique
8 semaines 1 à 2 heures par jour |
Amélioration des capacités de lecture
Augmentation de l'activation dans le gyrus temporal supérieur et la région pariétale inférieure gauche |
|
Temple et coll., 2003
|
IRMf
|
20 Dys (9,9)
12 NL (10,7) |
Stimuli non verbaux et stimuli de parole ralentis
8 semaines 100 mn par jour 5 jours par semaine |
Amélioration des capacités de langage oral et des performances de lecture
Augmentation de l'activation au niveau du cortex temporo-pariétal gauche et du gyrus frontal inférieur gauche |
|
sIRM : IRM spectroscopique ; PEs : potentiels evoqués ; MMN : mismatch negativity MEG : magneto-encéphalographique ;
|
||||
Un des rôles possibles de l'imagerie fonctionnelle pourrait être de permettre un diagnostic précoce de l'enfant dyslexique, afin de débuter des traitements avant son entrée dans l'apprentissage de la lecture. L'amélioration du dépistage des enfants à risque de développer une dyslexie peut permettre de mettre en place des entraînements précoces afin de parvenir à une réorganisation cérébrale optimale pour améliorer le trouble de la lecture (Gallagher et coll., 2000 ; Pennington et Lefly, 2001). L'évaluation comportementale des enfants d'âge préscolaire constitue sans doute la méthode de dépistage la plus efficace d'un point de vue économique (Fawcett et coll., 2003). Toutefois, la neuro-imagerie pourrait contribuer au diagnostic précoce dans des études familiales où les données génétiques et neuropsychologiques permettent d'individualiser des sujets à risque. Simos et coll. (2002b) retrouvaient chez des enfants âgés entre 5 et 6 ans, un pattern spécifique d'activation cérébrale caractérisé par une diminution de l'activité dans la région temporo-pariétale gauche et une augmentation de l'activité au niveau de la région homolatérale lors d'une tâche de reconnaissance de lettre. Ce résultat converge avec ceux obtenus chez des enfants considérés comme à risque de présenter une dyslexie du fait des antécédents familiaux, et chez lesquels les PE étaient prédictifs de la survenue d'une dyslexie (Leppanen et coll., 1999). Une interrogation d'ordre éthique doit être suscitée vis-à-vis du bien-fondé de ce type de dépistage dans la mesure où une mauvaise maîtrise des pratiques pourrait aboutir à inquiéter les familles et à stigmatiser les enfants concernés sans que les moyens de prise en charge rendus nécessaire par l'identification précoce du handicap ne soient correctement mis en œuvre.
; Pennington et Lefly, 2001). L'évaluation comportementale des enfants d'âge préscolaire constitue sans doute la méthode de dépistage la plus efficace d'un point de vue économique (Fawcett et coll., 2003). Toutefois, la neuro-imagerie pourrait contribuer au diagnostic précoce dans des études familiales où les données génétiques et neuropsychologiques permettent d'individualiser des sujets à risque. Simos et coll. (2002b) retrouvaient chez des enfants âgés entre 5 et 6 ans, un pattern spécifique d'activation cérébrale caractérisé par une diminution de l'activité dans la région temporo-pariétale gauche et une augmentation de l'activité au niveau de la région homolatérale lors d'une tâche de reconnaissance de lettre. Ce résultat converge avec ceux obtenus chez des enfants considérés comme à risque de présenter une dyslexie du fait des antécédents familiaux, et chez lesquels les PE étaient prédictifs de la survenue d'une dyslexie (Leppanen et coll., 1999). Une interrogation d'ordre éthique doit être suscitée vis-à-vis du bien-fondé de ce type de dépistage dans la mesure où une mauvaise maîtrise des pratiques pourrait aboutir à inquiéter les familles et à stigmatiser les enfants concernés sans que les moyens de prise en charge rendus nécessaire par l'identification précoce du handicap ne soient correctement mis en œuvre.En conclusion,
bien que les controverses continuent concernant la physiopathologie ou le traitement de la dyslexie, les différentes hypothèses pourraient être envisagées comme complémentaires plutôt que de s'exclure mutuellement. Les résultats des études en neuro-imagerie pourraient encourager la vérification d'hypothèses spécifiques. Les études d'imagerie fonctionnelle montrent non seulement des activations et une connectivité anormale des régions postérieures et péri-sylviennes gauches, mais également l'existence d'une grande plasticité cérébrale à la suite de divers types d'interventions. Toutefois, la spécificité des changements au niveau cognitif et neuronal observés au cours du développement, soit du fait de mécanismes de compensation, soit sous l'effet de traitement, reste à déterminer.
Les futures études concernant la physiopathologie des dyslexies de développement devront inclure des études comportementales et des études en imagerie fonctionnelle dans de larges séries longitudinales de sujets dyslexiques de nationalités différentes en s'intéressant non seulement à la lecture de mots isolés (comme la plupart des études actuellement) mais également aux domaines cognitifs plus complexes impliqués dans la lecture d'un texte. Enfin, les recherches sur les aspects thérapeutiques devront développer des programmes d'intervention intensive mais aussi spécifique, sa nature étant définie en fonction du profil neuropsychologique observé chez chaque patient et à une période déterminée de son évolution.
Bibliographie
[1] amitay s, ben-yehudah g, banai k, ahissar m. Disabled readers suffer from visual and auditory impairments but not from a specific magnocellular deficit.
Brain. 2002;
125:2272- 2285
[2] baldeweg t, richardson a, watkins s, foale c, gruzelier j. Impaired auditory frequency discrimination in dyslexia detected with mismatch evoked potentials.
Ann Neurol. 1999;
45:495- 503
[3] beaulieu c, plewes c, paulson la, roy d, snook l. et coll. Imaging brain connectivity in children with diverse reading ability.
NeuroImage. 2005;
25:1266- 1271
[4] bednarek db, grabowska a. Luminance and chromatic contrast sensitivity in dyslexia: the magnocellular deficit hypothesis revisited.
Neuroreport. 2002;
13:2521- 2525
[5] breznitz z, leikin m. Syntactic processing of Hebrew sentences in normal and dyslexic readers: electrophysiological evidence.
J Genet Psychol. 2000;
161:359- 380
[6] brunswick n, mccrory e, price cj, frith cd, frith u. Explicit and implicit processing of words and pseudowords by adult developmental dyslexics: A search for Wernicke’s Wortschatz?.
Brain. 1999;
122:1901- 1917
[7] cohen l, lehericy s, chochon f, lemer c, rivaud s, dehaene s. Language-specific tuning of visual cortex? Functional properties of the Visual Word Form Area.
Brain. 2002;
125:1054- 1069
[8] corina dp, richards tl, serafini s, richards al, steury k. et coll. fMRI auditory language differences between dyslexic and able reading children.
Neuroreport. 2001;
12:1195- 1201
[9] démonet jf, fiez ja, paulesu e, petersen se, zatorre rj. PET studies of phonological processing. A critical reply to Poeppel.
Brain Lang. 1996;
28:352- 385
[10] démonet jf, thierry g, cardebat d. Renewal of the neurophysiology of language: functional neuroimaging.
Physiol Rev. 2005;
85:49- 95
[11] deutsch gk, dougherty rf, bammer r, siok wt, gabrieli jde, wandell b. Children’s reading performance is correlated with white matter structure measured by diffusion tensor imaging.
Cortex. 2005;
41:354- 363
[12] eckert ma, leonard cm, richards tl, aylward e, thomson j, berninger v. Anatomical correlates of dyslexia: frontal and cerebellar findings.
Brain. 2003;
126:482- 494
[13] eden gf, vanmeter jw, rumsey jm, maisog jm, woods rp, zeffiro ta. Abnormal processing of visual motion in dyslexia revealed by functional brain imaging.
Nature. 1996;
382:66- 69
[14] eden gf, jones km, cappell k, gareau l, wood fbet coll.. Clinical Study Neural Changes following Remediation in Adult Developmental Dyslexia.
Neuron. 2004;
44:411- 422
[15] fawcett aj, nicolson ri, lee r. The pre-school screening test.
London:Psychological Corporation;
2003.
[16] galaburda am, kemper tl. Cytoarchitectonic abnormalities in developmental dyslexia: a case study.
Ann Neurol. 1979;
6:94- 100
[17] galaburda am, sherman gf, rosen gd, aboiti f, geschwind n. Developmental dyslexia: four consecutive patients with cortical anomalies.
Ann Neurol. 1985;
18:222- 233
[18] gallagher am, frith u, snowling m. Precursors of literacy delay among children at genetic risk of dyslexia.
J Child Psych Psychia. 2000;
41:202- 213
[19] georgiewa p, rzanny r, hopf jm, knab r, glauche v. et coll. fMRI during word processing in dyslexic and normal reading children.
Neuroreport. 1999;
10:3459- 3465
[20] georgiewa p, rzanny r, gaser c, gerhard uj, vieweg u. et coll. Phonological processing in dyslexic children: A study combining functional imaging and event related potentials.
Neurosci Lett. 2002;
318:5- 8
[21] giraud k, démonet jf, habib m, marquis p, chauvel p, liegeois-chauvel c. Auditory Evoked Potential Patterns to Voiced and Voiceless Speech Sounds in Adult Developmental Dyslexics with Persistent Deficits.
Cereb Cortex. 2005;
15:1524- 1534
[22] habib m. The neurological basis of developmental dyslexia: an overview and working hypothesis.
Brain. 2000;
123:2373- 2399
[23] hatcher p, hulme c. Phonemes, rhymes, and intelligence as predictors of children’s responsiveness to remedial reading instruction: Evidence from a longitudinal intervention study.
J Exp Child Psych. 1999;
72:130- 153
[24] helenius p, tarkiainen a, cornelissen p, hansen pc, salmelin r. Dissociation of normal feature analysis and deficient processing of letter-strings in dyslexic adults.
Cereb Cortex. 1999;
9:476- 483
[25] horwitz b, rumsey jm, donohue bc. Functional connectivity of the angular gyrus in normal reading and dyslexia.
Proc Natl Acad Sci USA. 1998;
95:8939- 8944
[26] jenner ar, galaburda am, sherman gf. Connectivity of ectopic neurons in the molecular layer of the somatosensory cortex in autoimmune mice.
Cerebral Cortex. 2000;
10:1005- 1013
[27] johannes s, kussmaul cl, munte tf, mangun gr. Developmental dyslexia: passive visual stimulation provides no evidence for a magnocellular processing defect.
Neuropsychologia. 1996;
34:1123- 1127
[28] khan sc, frisk v, taylor mj. Neurophysiological measures of reading difficulty in very-low- birthweight children.
Psychophysiology. 1999;
36:76- 85
[29] klingberg t, hedehus m, temple e, salz t, gabrieli jd. et coll. Microstructure of temporo-parietal white matter as a basis for reading ability: evidence from diffusion tensor magnetic resonance imaging.
Neuron. 2000;
25:493- 500
[30] kraus n, mcgee tj, carrell td, zecker sg, nicol tg, koch db. Auditory neurophysiologic responses and discrimination deficits in children with learning problems.
Science. 1996;
273:971- 973
[31] kujala t, karma k, ceponiene r, belitz s, turkkila p. et al. Plastic neural changes and reading improvement caused by audiovisual training in reading-impaired children.
Proc Natl Acad Sci USA. 2001;
98:10509- 10514
[32] kujala t, naatanen r. The mismatch negativity in evaluating central auditory dysfunction in dyslexia.
Neurosci Biobehav Rev. 2001;
25:535- 543
[33] leonard cm, eckert ma, lombardino lj, oakland t, kranzler j. et coll. Anatomical risk factors for phonological dyslexia.
Cereb Cortex. 2001;
11:148- 157
[34] leppanen ph, pihko e, eklund km, lyytinen h. Cortical responses of infants with and without a genetic risk for dyslexia: II. Group effects.
Neuroreport. 1999;
10:969- 973
[35] livingstone ms, rosen gd, drislane fw, galaburda am. Physiological and anatomical evidence for a magnocellular defect in developmental dyslexia.
Proc Natl Acad Sci USA. 1991;
88:7943- 7947
[36] nagarajan s, mahncke h, salz t, tallal p, roberts t, merzenich mm. Cortical auditory signal processing in poor readers.
Proc Natl Acad Sci USA. 1999;
96:6483- 6488
[37] nicolson ri, fawcett aj, berry el, jenkins ih, dean p, brooks dj. Association of abnormal cerebellar activation with motor learning difficulties in dyslexic adults.
Lancet. 1999;
353:1662- 1667
[38] paulesu e, frith u, snowling m, gallagher a, morton j. et coll. Is developmental dyslexia a disconnection syndrome? Evidence from PET scanning.
Brain. 1996;
119:143- 157
[39] paulesu e, démonet jf, fazio f, mccrory e, chanoine v. et coll. Dyslexia: cultural diversity and biological unity.
Science. 2001;
291:2165- 2167
[40] pennington bf, lefly dl. Early reading development in children at family risk for dyslexia.
Child Devel. 2001;
72:816- 833
[41] price cj. The functional anatomy of word comprehension and production.
Trends Cogn Sci. 1998;
2:281- 288
[42] price cj, warburton ea, moore cj, frackowiak rs, friston kj. Dynamic diaschisis: anatomically remote and context-sensitive human brain lesions.
J Cogn Neurosci. 2001;
13:419- 429
[43] pugh kr, mencl we, jenner ar, katz l, frost sjet coll.. Functional neuroimaging studies of reading and reading disability (developmental dyslexia).
Ment Retard Dev Disabil Res Rev. 2000;
6:207- 213
[44] rae c, lee ma, dixon rm, blamire am, thompson ch. et coll. Metabolic abnormalities in developmental dyslexia detected by 1H magnetic resonance spectroscopy.
Lancet. 1998;
351:1849- 1852
[45] rae c, harasty ja, dzendrowskyj te, talcott jb, simpson jmet coll.. Cerebellar morphology in developmental dyslexia.
Neuropsychologia. 2002;
40:1285- 1292
[46] richards tl, dager sr, corina d, serafini s, heide acet coll.. Dyslexic children have abnormal brain lactate response to reading-related language tasks.
AmJ Neuroradiol. 1999;
20:1393- 1398
[47] richards tl, corina d, serafini s, steury k, echelard dr. et coll. Effects of a phonologically driven treatment for dyslexia on lactate levels measured by proton MR spectroscopic imaging.
Am J Neuroradiol. 2000;
21:916- 922
[48] ruff s, cardebat d, marie n, démonet jf. Enhanced response of the left frontal cortex to slowed down speech in dyslexia: an fMRI study.
Neuroreport. 2002;
13:1285- 1289
[49] rumsey jm, andreason p, zametkin aj, aquino t, king ac. et al. Failure to activate the left temporoparietal cortex in dyslexia. An oxygen 15 positron emission tomographic study.
Arch Neurol. 1992;
49:527- 534
[50] rumsey jm, donohue bc, brady dr, nace k, giedd jn, andreason p. A magnetic resonance imaging study of planum temporale asymmetry in men with developmental dyslexia.
Arch Neurol. 1997;
54:1481- 1489
[51] rumsey jm, horwitz b, donohue bc, nace kl, maisog jm, andreason pa. Functional lesion in developmental dyslexia: left angular gyral blood flow predicts severity.
Brain Lang. 1999;
70:187- 204
[52] schulte-körne g, deimel w, bartling j, remschmidt h. Auditory processing and dyslexia: evidence for specific speech processing deficit.
NeuroReport. 1998;
9:337- 340
[53] shaywitz se. Dyslexia.
N Engl J Med. 1998;
338:307- 312
[54] shaywitz se, shaywitz ba, pugh kr, fulbright rk, constable rt. et coll. Functional disruption in the organization of the brain for reading in dyslexia.
Proc Natl Acad Sci USA. 1998;
95:2636- 2641
[55] shaywitz se, fletcher jm, holahan jm, shneider ae, marchione ke. et coll. Persistance of dyslexia: the Connecticut Longitudinal Study at adolescence.
Pediatrics. 1999;
104:1351- 1359
[56] shaywitz ba, shaywitz se, pugh kr, mencl we, fulbright rk. et coll. Disruption of posterior brain systems for reading in children with developmental dyslexia.
Biol Psychiatry. 2002;
52:101- 110
[57] shaywitz ba, shaywitz se, blachman b, pugh kr, fulbright rk. et coll. Development of Left Occipitotemporal Systems for Skilled Reading in Children After a Phonologically Based Intervention.
Biol Psychiatry. 2004 doi:10.1016/j.biopsych.2003.12.019;
:-
[58] shaywitz se, shaywitz ba, fulbright rk, skudlarski p, mencl weet coll.. Neural systems for compensation and persistence : young adult outcome of childhood reading disability.
Biol Psychiatry. 2003;
54:25- 33
[59] silani g, frith u, démonet jf, fazio f, perani d. et coll. Brain abnormalities underlying altered activation in dyslexia: a voxel based morphometry study.
Brain. 2005;
128:2453- 2461
[60] simos pg, breier ji, fletcher jm, bergman e, papanicolaou ac. Cerebral mechanisms involved in word reading in dyslexic children: a magnetic source imaging approach.
Cereb Cortex. 2000a;
10:809- 816
[61] simos pg, breier ji, fletcher jm, maggio ww, fletcher jmet coll.. Brain activation in dyslexic in dyslexic children during non-word reading: a magnetic source imaging study.
Cereb Neurosci Letters. 2000b;
290:61- 65
[62] simos pg, breier ji, wheless jw, maggio ww, fletcher jmet coll.. Brain mechanisms for reading : the role of the superior temporal gyrus in word and pseudoword naming.
Neuroreport. 2000c;
11:2443- 2467
[63] simos pg, fletcher jm, bergman e, breier ji, foorman bret coll.. Dyslexia-specific brain activation profile becomes normal following successful remedial training.
Neurology. 2002a;
58:1203- 1213
[64] simos pg, fletcher jm, foorman br, francis dj, castillo em. et coll. Brain activation profiles during the early stages of reading acquisition.
J Child Neuro. 2002b;
17:159- 163
[65] taylor mj, keenan nk. Event-related potentials to visual and language stimuli in normal and dyslexic children.
Psychophysiology. 1990;
27:318- 327
[66] taylor mj, keenan nk. ERPs to orthographic, phonological and semantic classification tasks in normal and dyslexic children.
Devel Neuropsych. 1999;
15:307- 326
[67] temple e, poldrack ra, salidis j, deutsch gk, tallal p. et coll. Disrupted neural responses to phonological and orthographic processing in dyslexic children: an fMRI study.
Neuroreport. 2001;
12:299- 307
[68] temple e, deutsch gk, poldrack ra, miller sl, tallal p. et coll. Neural deficits in children with dyslexia ameliorated by behavioral remediation: evidence from functional MRI.
Proc Natl Acad Sci USA. 2003;
100:2860- 2865
[69] torgensen jk, wagner rk, rashotte ca, rose e, lindamood p. et coll. Preventing reading failure in young children with phonological processing disabilities: group and individual responses to instruction.
J Educ Psychol. 1999;
91:579- 593
[70] vanni s, uusitalo ma, kiesila p, hari r. Visual motion activates V5 in dyslexics.
Neuroreport. 1997;
8:1939- 1942
[71] wise bw, ring j, olson rk. Individual differences in gains from computer-assisted remedial reading.
J Exp Child Psych. 2000;
77:197- 335
→ Aller vers SYNTHESE