| |
| Med Sci (Paris). 2014 October; 30(10): 902–909. Published online 2014 October 14. doi: 10.1051/medsci/20143010018.La lumière comme stimulant de l’activité cognitive cérébrale Gilles Vandewalle1* 1Université de Liège, centre de recherches du cyclotron, bâtiment B30, 4000Liège, Belgique |
La lumière est nécessaire à la vision, mais joue aussi un rôle essentiel pour de nombreuses fonctions non visuelles (Figure 1). L’exposition lumineuse est en effet le synchronisateur principal de l’horloge circadienne sensible à l’alternance jour-nuit, ce qui permet de réguler le comportement et la physiologie de façon optimale au cours d’une journée de 24 h [
1]. La lumière affecte ainsi la rythmicité circadienne d’une multitude de fonctions telles que la thermorégulation, la sécrétion de certaines hormones (mélatonine, cortisol), l’alternance veille-sommeil, le niveau d’éveil et les performances cognitives [
35]. Le manque de lumière semble aussi jouer un rôle important pour l’humeur, et la photothérapie (luminothérapie) est recommandée pour traiter la dépression saisonnière [
2,
36]. En plus de ces effets à long terme, la lumière a également des effets directs sur des fonctions aussi variées que le rythme cardiaque, la taille pupillaire, la sécrétion de cortisol et de mélatonine, ainsi qu’un impact stimulant sur l’éveil et la cognition [
3] ; ce dernier impact stimulant est très important. Cette revue va se concentrer sur cet aspect.
 | Figure 1.
Étendue des influences non visuelles de la lumière. Un réseau diffus de cellules ganglionnaires rétiniennes (CGR) intrinsèquement photosensibles (p) grâce à l’expression de la mélanopsine, est plus sensible au bleu (480 nm) et reçoit des afférences des cônes et des bâtonnets (A). Ces cellules ont des projections directes vers les noyaux suprachiasmatiques (SCN) de l’hypothalamus, site de l’horloge circadienne principale, et régulent l’entraînement circadien, notamment via une influence sur l’expression de certains gènes (B). La lumière régule également l’activité locomotrice et peut modifier la phase circadienne (C). Les CGR à mélanopsine se connectent au prétectum contrôlant la constriction pupillaire (D), et des projections indirectes via le SCN sont responsables de la suppression de la sécrétion de mélatonine par la lumière (E). Les CGR à mélanopsine ont aussi des projections directes vers des structures régulant le sommeil, telles que l’aire ventrolatérale préoptique (VLPO) et affectent donc le sommeil (F). La lumière bleue modifie les réponses cognitives cérébrales (G) et peut améliorer le niveau d’éveil et de vigilance (H). Les CGR à mélanopsine sont aussi impliquées dans des fonctions visuelles rudimentaires (Figure extraite de [
33]). |
Il a été démontré qu’une exposition lumineuse augmentait directement les performances pour des tâches mettant en jeu des processus exécutifs, tels qu’inhibition, changement de tâche (shifting), mise à jour, processus mnésiques incluant mémoire de travail et mémoire à long terme, et processus attentionnels [
4]. La lumière réduit aussi directement la puissance spectrale à l’éveil des bandes de fréquences delta (0,5-4 Hz), thêta (4-8 Hz) et alpha (8-12 Hz) de l’électroencéphalogramme (EEG), ainsi que les mouvements oculaires lents, qui sont tous des marqueurs de somnolence et de déficits attentionnels [
5]. Ces derniers résultats constituent en fait les premiers éléments des mécanismes cérébraux responsables de l’effet stimulant de la lumière. Au cours des dix dernières années, mes collègues et moi-même, à Liège, Montréal et dans le Surrey (Royaume-Uni), avons réalisé une série d’expériences de neuro-imagerie qui ont élucidé une partie de ces processus cérébraux. Cet article synthétise ces études. |
Large impact cérébral de la lumière Lors de nos études, les participants effectuaient des tâches cognitives pendant qu’ils étaient, soit maintenus dans l’obscurité, soit éclairés. Nous avons caractérisé l’impact de la lumière sur l’activité cérébrale mise en place pour réaliser la tâche [4]. En d’autres termes, la lumière n’était pas un aspect important de la tâche pour le participant et servait à manipuler le contexte dans lequel la tâche était effectuée. Nous avons par ailleurs pris soin d’utiliser des protocoles qui dépendaient le moins possible des fonctions visuelles. Les tâches utilisées étaient en effet effectuées dans différentes versions auditives. La durée d’illumination était relativement longue (17 à 20 min) dans un premier temps, puis beaucoup plus courtes dans un second temps (40 à 60 s). La lumière était administrée via des surfaces diffuses qui occupaient tout le champ visuel (le système visuel est surtout sensible aux changements et aux contrastes). Nos résultats démontrent que la lumière a un impact très important sur l’activité cérébrale qui sous-tend l’attention, seule (paradigme oddball) ou en combinaison avec des processus de mise à jour et de comparaison (tâche de type n-back), la mémoire de travail (n-back), ou le traitement de stimulations émotionnelles [4,
6] (Figure 2). Ce dernier aspect émotionnel étend les influences cognitives connues de la lumière et ouvre peut-être de nouvelles perspectives pour les traitements lumineux des troubles de l’humeur à long terme.
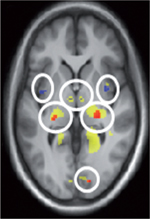
 | Figure 2.
Localisations des effets de la lumière sur l’activité cognitive cérébrale non visuelle. Des effets non visuels de la lumière ont été observés durant une exposition monochromatique bleue, ou juste après une exposition à la lumière blanche très intense, notamment dans les régions suivantes (de A à H, de gauche à droite et de haut en bas) : hypothalamus (A) dans des zones incluant le SCN, le VLPO, le DMH ou le PVNh ; tronc cérébral dans une région comprenant le locus cœruleus
(B) ; thalamus dorso-postérieur (pulvinar) (C) ; amygdale (D) ; hippocampe (E) ; cortex préfrontal (F) ; dans l’insula (G) ; dans le cortex pariétal (H) (droite : sillon intrapariétal ; gauche : gyrus supramarginal) (d’après [ 4] avec autorisation). |
Nos études ont mis en évidence que la lumière modulait rapidement l’activité de régions sous-corticales impliquées dans la régulation de l’éveil [4,
7,
8]. Ces régions comprennent tout d’abord l’hypothalamus. L’imagerie par résonance magnétique ne permet pas d’établir avec certitude quels noyaux sont affectés mais, pour des tâches attentionnelles, il pourrait s’agir des noyaux suprachiasmatiques (SCN, site de l’horloge circadienne), des noyaux ventrolatéraux pré-optiques (VLPO, qui régulent le sommeil et l’éveil) [
9] ou, pour des tâches mettant en jeu les émotions, des noyaux hypothalamiques dorsomédians (DMH) ou paraventriculaires (PVNh), tous deux impliqués dans les réponses émotionnelles [6]. La lumière semble aussi avoir un effet très rapide au niveau du tronc cérébral, probablement au niveau des noyaux de la formation réticulée constituant le système activateur ascendant, tels que le locus cœruleus (LC), source principale de noradrénaline, et le raphé dorsal (DR), source de sérotonine [8]. Tout comme l’hypothalamus, ces noyaux ont des projections monosynaptiques vers toutes les zones du cortex et pourraient facilement répercuter l’influence de la lumière dans tout le cerveau [
10]. Par ailleurs, le pulvinar du thalamus, qui est impliqué dans la régulation de l’éveil, de l’attention et des ressources cognitives, est la structure la plus souvent affectée par la lumière dans nos études et semble jouer un rôle essentiel dans l’impact non visuel de la lumière sur la cognition [4, 7, 8]. Un impact de la lumière sur le flux d’informations des boucles thalamocorticales pourrait affecter de larges territoires corticaux. Enfin, l’activité de l’hippocampe et de l’amygdale, respectivement impliquées dans la mémoire à long terme et dans la régulation des émotions, est aussi affectée par la lumière [6,
11,
12]. Au niveau cortical, nos résultats montrent que l’impact de la lumière dépend du processus cognitif en cours. Une tâche plus attentionnelle mettra en évidence des modulations au niveau du cortex dorsolatéral préfrontal et du sillon intrapariétal [9, 12]. Une tâche de mémoire de travail, mettant en jeu des fonctions exécutives (comparaison, mise à jour) en plus de l’attention, verra l’activité de régions pariétales supérieures impliquées dans ces processus, ainsi que celle des régions fronto-polaires et ventrolatérales du cortex préfrontal affectées par la lumière [7, 8, 11,
13]. Il est important de souligner que ces régions préfrontales sont considérées actuellement comme au sommet de la hiérarchie cognitive et engagées dans les processus les plus complexes [
14,
37], soulignant l’importance de la lumière pour les fonctions cognitives supérieures. Le niveau d’activité des cortex sensoriels occipitaux ou temporaux est aussi influencé par la lumière, probablement via une influence top-down des régions pariétales ou préfrontales [9, 12]. Une tâche mettant en jeu les émotions et, dans ce cas précis, le processing de stimulations vocales émotionnelles (dépourvues de signification) a permis de mettre en évidence un impact de la lumière au niveau de « l’aire de la voix » dans le cortex temporal, qui est impliquée dans le décodage de l’information émotionnelle vocale [6]. En résumé, nos études montrent que la lumière exerce une large influence sur le fonctionnement cérébral, qu’il s’agisse de régions sous-corticales régulant le niveau d’éveil ou de régions corticales qui jouent des rôles centraux dans la régulation de l’attention, des émotions et des fonctions exécutives supérieures. |
L’impact de la lumière dépend de sa longueur d’onde et de la mélanopsine Nos premières études ont utilisé une lumière blanche (qui contient toutes les longueurs d’onde) pour démontrer un impact persistant de la lumière dans la période d’obscurité qui suivait immédiatement l’exposition [9, 12]. Nous avons ensuite démontré que, durant l’exposition lumineuse, les courtes longueurs d’onde étaient plus efficaces que les longues longueurs d’onde pour augmenter l’activité cérébrale liée à un processus cognitif en cours [6–8, 13]. Ainsi, par rapport à une lumière monochromatique verte (525 ou 550 nm), une lumière bleue (460 ou 480 nm) augmentait les niveaux d’activité du pulvinar, ainsi que celui des régions préfrontales et pariétales impliquées dans une tâche de mémoire de travail, ou bien celui de l’hypothalamus, de l’amygdale, de l’hippocampe et de la région temporale de la voix lors d’une tâche auditive émotionnelle. Cette supériorité de la lumière bleue est en accord avec la littérature qui montre que l’amélioration des performances cognitives est plus importante avec des lumières bleues ou enrichies en bleu [3,
15,
16]. Elle est due à un type de photorécepteur rétinien qui n’a été découvert que très récemment : les cellules ganglionnaires rétiniennes (CGR), qui sont photosensibles parce qu’elles expriment la mélanopsine, un photopigment plus sensible au bleu [
17,
18]. Ces CGR reçoivent des afférences des cônes et des bâtonnets, qui s’ajoutent à leur propre photosensibilité, et se projettent en de multiples sites sous-corticaux responsables des effets non visuels de la lumière. Ces afférences sont responsables du rôle important non visuel de la lumière. Le rôle des CGR à mélanopsine dans les fonctions non visuelles de la lumière est bien établi dans des modèles animaux [17, 18]. Ces modèles animaux sont cependant principalement des rongeurs nocturnes incapables de processus cognitifs complexes. Nos recherches, ainsi que celles d’autres chercheurs, sur l’impact de la lumière sur la cognition et l’éveil, sont donc essentielles. Les manipulations moléculaires ou génétiques réalisées chez le rongeur pour séparer cet impact non visuel de l’influence des cônes et des bâtonnets sont impossibles chez l’homme. Les preuves d’un rôle de la mélanopsine pour la cognition sont restées principalement indirectes (bien que le bleu ait un impact plus important, cela n’exclut pas que les cônes ou les bâtonnets en soient exclusivement ou en partie responsables) [3, 15, 16]. Nous avons réalisé deux études pour tenter d’apporter des preuves plus directes de l’implication des cellules ganglionnaires rétiniennes à mélanopsine. Nous avons tout d’abord étudié des personnes totalement aveugles chez lesquelles la lumière produit encore des effets non visuels (contraction de la pupille, suppression de la mélatonine), très probablement parce qu’elles conservent les CGR à mélanopsine [
19]. Notre étude démontre que, malgré le fait que ces individus ne voient absolument rien, de courtes expositions à la lumière bleue intense ont une grande influence sur l’activité cérébrale mise en place pour réaliser une tâche auditive de mémoire de travail, au niveau du pulvinar, et des cortex préfrontal et occipital (Figure 3). Au moins une partie de l’impact de la lumière sur les fonctions cognitives non visuelles semble donc être due aux CGR à mélanopsine. Cependant, ces aveugles sensibles à la lumière sont très rares (neuf personnes identifiées dans le monde).
 | Figure 3.
Impact de la lumière chez des personnes totalement aveugles, mais conservant des réponses non visuelles à la lumière (suppression de la mélatonine, contraction pupillaire). Par rapport à l’obscurité totale, une lumière bleue intense augmente nettement les réponses cérébrales mises en place pour la réalisation d’une tâche de mémoire de travail chez des personnes totalement aveugles, qui sont donc incapables de dire si la lumière est allumée ou pas. Ces effets ont été détectés dans le cortex occipital (A), dans le pulvinar (gauche) et le cortex préfrontal médian (droite) (B), ainsi que dans le cortex préfrontal latéral (C), important pour la régulation de l’éveil et des fonctions exécutives supérieures. D. Estimation de l’activité cérébrale enregistrée dans ces régions sous lumière bleue ou dans l’obscurité (adapté de [ 19] avec autorisation). |
Il restait à établir l’implication des CGR à mélanopsine chez des personnes aux fonctions visuelles normales. Nous avons démontré que chez ces individus, l’influence d’une lumière donnée sur l’activité cérébrale nécessaire à une tâche auditive de mémoire de travail dépendait de la longueur d’onde de la lumière reçue précédemment [11]. Notre protocole était conçu de telle sorte que les cônes et les bâtonnets ne pouvaient être responsables de cet effet, et seules les propriétés de la mélanopsine pouvaient l’expliquer. La mélanopsine a en effet des caractéristiques identiques à celles des photopigments d’invertébrés dont la sensibilité est influencée par la lumière reçue précédemment [
20]. Ainsi, alors que les personnes que nous enregistrions étaient placées dans des conditions strictement identiques, l’impact d’une lumière test sur l’activité du pulvinar et du cortex préfrontal était augmenté si une exposition à une lumière orange précédait l’enregistrement (de plus d’une heure, avec une période d’obscurité totale entre la lumière orange et la lumière test). Cette dernière étude constitue la démonstration la plus claire à ce jour d’un rôle dans la cognition de la mélanopsine et des CGR qui sont exprimés chez des individus aux fonctions visuelles normales [11]. L’influence de l’historique lumineux pourrait donner à la mélanopsine un rôle de « mémoire lumineuse » pour la cognition, en intégrant l’exposition lumineuse sur de longues périodes de temps. |
L’influence de la lumière change avec sa durée et son intensité, mais aussi avec le manque de sommeil et le moment de la journée Lorsque la lumière est reçue pendant des durées plus courtes (environ 1 min), son impact semble principalement moduler l’activité des régions sous-corticales [4]. Si la durée augmente (16-20 min), un impact prononcé au niveau cortical est détecté dans les régions impliquées dans le processus cognitif en cours [4]. Nos résultats suggèrent enfin que si cet impact cortical est suffisamment fort lors de l’exposition à des lumières très intenses de plus longue durée (20 min), un effet de la lumière peut être détecté de façon significative dans des mesures comportementales [9, 12]. Nos résultats pointent vers un élargissement graduel de l’impact de la lumière avec sa durée et son intensité, qui se traduit progressivement par un effet comportemental. En accord avec ces propositions, d’autres études utilisant uniquement des variables comportementales ou électroencéphalographiques (EEG) ont rapporté un impact significatif au bout de 30 min d’illumination avec des lumières bleues, ou enrichies en bleu, de longueur d’onde et d’intensité comparables à celles que nous avons employées [15, 16,
21,
22]. La durée, l’intensité et la longueur d’onde ne sont cependant pas les seuls aspects à prendre en considération lors de l’administration d’une exposition lumineuse. Nous avons en effet étudié l’impact non visuel de la lumière sur les réponses cognitives cérébrales à différents moments de la journée et lors d’une privation de sommeil [7]. Il ressort de nos recherches que l’impact de la lumière bleue dépend des mécanismes endogènes déjà en place pour réguler l’éveil. Le matin, peu de temps après le réveil, la lumière bleue semble avoir un impact sur les réponses corticales liées à la tâche en cours. En soirée, lorsque le système circadien (horloge biologique) envoie un signal fort pour promouvoir l’éveil, l’impact de la lumière est diminué [7]. La lumière semble donc moins capable « d’aider » les réponses cérébrales dans les conditions où l’éveil est déjà stimulé par des mécanismes endogènes. À l’inverse, le matin qui suit une nuit de privation de sommeil, lorsque notre système lutte difficilement contre le manque de sommeil, la lumière bleue a un impact étendu tant au niveau cortical qu’au niveau du pulvinar [7]. |
Marqueurs de différences interindividuelles dans l’impact de la lumière : génétique, âge et santé mentale Nos recherches se sont aussi focalisées sur les marqueurs individuels qui pouvaient modifier l’impact de la lumière. Nous avons d’abord montré que le polymorphisme du gène PERIOD3, qui affecte la régulation du sommeil et de l’éveil [
23], augmentait ou diminuait l’impact de la lumière sur les fonctions cognitives cérébrales en fonction du moment de la journée. Les individus homozygotes pour la forme longue du gène (PER3
5/5
), qui sont plus affectés par le manque de sommeil que les individus homozygotes pour sa forme courte (PER3
4/4
), montrent un effet plus grand de la lumière bleue sur leur réponse cérébrale [7]. Il est donc possible que la lumière ait un impact plus important sur les individus en plus grande difficulté face au manque de sommeil, et qu’elle puisse aider à « sauver » leur activité cérébrale. Ces résultats renforcent également l’idée selon laquelle l’impact de la lumière dépend des mécanismes endogènes de régulation de l’éveil. L’âge affecte profondément notre physiologie, notre sommeil et nos rythmes biologiques [
24], et plusieurs recherches suggèrent que l’impact de la lumière sur certaines fonctions non visuelles diminue avec l’âge [
25]. Puisque cette diminution ne semble pas être observée pour toutes les fonctions non visuelles [
26,
27], il était important de savoir si l’impact cognitif de la lumière était préservé ou non. Notre première étude sur ces aspects indique que la lumière bleue est toujours capable de stimuler l’activité cérébrale chez des personnes plus âgées, et qu’on peut donc envisager d’utiliser la lumière comme aide cognitive aussi au cours du vieillissement [
28]. Nos résultats montrent cependant que l’impact de la lumière bleue sur l’activité cérébrale diminue avec l’âge dans plusieurs régions clés pour la cognition et l’éveil (cortex préfrontal, amygdale, pulvinar) [28] (Figure 4). Les caractéristiques optimales de l’environnement lumineux doivent donc peut-être être adaptées pour aider la cognition au cours du vieillissement. D’autres recherches avec des lumières de longueur d’onde ou d’intensité différentes sont encore nécessaires pour établir les caractéristiques de l’effet stimulant au cours du vieillissement.
 | Figure 4.
L’impact de la lumière persiste mais diminue avec l’âge. Des individus jeunes (22,8 ± 4,1 ans) et plus âgés (60,9 ± 4,5 ans) en bonne santé ont réalisé une tâche de mémoire de travail, soit dans l’obscurité, soit sous une exposition à une lumière bleue. Les réponses détectées chez les individus jeunes sont en jaune, chez les individus plus âgés en rouge, et les différences significatives entre groupes d’âge sont en bleu. A. Pulvinar. B. Thalamus (haut), amygdale (bas). C. Insula bilatéral (haut, latéraux), thalamus (centre), corps géniculés latéraux (milieu, latéraux), cortex occipital (bas, scissure calcarine). D. Cortex préfrontal (haut), insula (milieu), gyrus lingual (bas). E. Exemple de réponses significatives à la lumière, présentes dans les deux groupes d’âge. F. Exemple de différence significative dans la réponse à la lumière entre les deux groupes d’âge (les individus jeunes ont une réponse significativement plus grande que les individus âgés, et, en fait, dans ce type de régions, il n’y a de réponse significative que chez les jeunes, et pas chez les plus âgés) (adapté de [ 28] avec autorisation). |
Enfin, nous avons pu montrer que la réponse à la lumière d’individus souffrant de dépression saisonnière était anormale au niveau de l’hypothalamus postérieur lors de la réalisation d’une tâche émotionnelle auditive [
29]. Par rapport aux sujets contrôles, l’activité d’une zone compatible avec l’hypothalamus latéral ou le noyau paraventriculaire hypothalamique était augmentée sous lumière bleue et diminuée sous lumière verte lors du traitement de vocalisations émotionnelles dénuées de sens. L’hypothalamus étant une région clé pour les rythmes biologiques, le sommeil, l’éveil et les émotions, ce résultat est intéressant. Il reste cependant à déterminer si cette réponse anormale, qui était présente durant la période symptomatique (automne-hiver), est aussi détectée durant la période de rémission (printemps-été), et si elle est responsable de l’impact du manque de lumière en hiver chez ces patients et de l’effet positif de la photothérapie sur leurs symptômes [37]. |
Conclusions : la lumière pour optimiser la cognition Nos recherches démontrent que la lumière joue un rôle prépondérant dans la régulation des fonctions cognitives cérébrales. Nous avons pu établir une partie des mécanismes cérébraux responsables de l’effet stimulant de la lumière démontrés par d’autres, en utilisant des tâches très diverses (Figure 5). Nous pensons que la lumière touche d’abord des régions sous-corticales impliquées dans la régulation de l’éveil avant d’affecter les régions corticales impliquées dans la tâche cognitive en cours. Cette modulation corticale dépend donc du type de processus recruté par cette tâche et, si l’effet de la lumière au niveau de cortex est suffisamment important, elle affectera les performances et le comportement.
 | Figure 5.
Schéma des mécanismes cérébraux impliqués dans l’impact de la lumière sur l’activité cognitive non visuelle. 1. À l’allumage, un impact de la lumière est observé dans l’hypothalamus (rond bleu foncé) et dans le pulvinar (ronds verts) (ainsi que dans l’amygdale et l’hippocampe, non représentés). 2. Durant les premières secondes de l’exposition, l’influence de la lumière est surtout détectée dans des structures sous-corticales régulant l’éveil et le sommeil (hypothalamus, tronc cérébral [rond jaune], pulvinar). 3. Des réponses plus tardives sont observées dans les régions corticales impliquées dans le processus cognitif en cours et peuvent affecter, par la suite, le comportement et les performances. Pour l’attention/mémoire de travail/tâches exécutives (ronds rouges), un réseau de régions centré sur le pulvinar et comprenant le cortex préfrontal et pariétal semble responsable de l’impact de la lumière sur l’éveil et la cognition. Pour les réponses à des stimulations émotionnelles vocales, le réseau implique l’hypothalamus, l’amygdale et la « région de la voix » du cortex temporal (ronds bleu clair). La lumière semble avoir un impact plus rapide sur les réponses émotionnelles que sur celles ayant trait à l’attention, à la mémoire de travail et aux réponses exécutives. L’impact de la lumière est plus important pour de plus fortes intensités, de plus longues durées et de plus courtes longueurs d’onde (bleues). Le moment de la journée, et les changements de pression de sommeil et de phase circadienne qui y sont associés modifient l’impact de la lumière, ainsi que l’âge, la santé mentale et le génotype (de PERIOD3) (adapté de [ 4] et [
34] avec autorisation). |
Nos recherches ne constituent pas un plaidoyer pour la lumière bleue. Tout comme d’autres travaux utilisant des lumières de différentes longueurs d’onde et de différentes intensités avec d’autres protocoles [15, 16, 21, 22], elles mettent en avant l’impact non visuel de la lumière. Elles renforcent l’idée selon laquelle il est important de considérer la lumière et ses fonctions non visuelles lorsqu’on s’expose à la lumière. Les lumières artificielles que nous utilisons sont généralement de faible intensité (< 300 lux) et sont souvent pauvres en longueurs d’onde bleues. Modifier ces éclairages pourrait avoir des effets très bénéfiques. Il a été rapporté, par exemple, qu’une fois enrichi en bleu, l’environnement lumineux améliore la perception subjective de la performance au travail et de la qualité du sommeil, ainsi que l’humeur [
30]. Comme autre exemple, augmenter l’intensité lumineuse dans les institutions pour patients âgés souffrants de démence ralentit la progression des symptômes, augmente l’activité le jour et la diminue la nuit, et améliore donc potentiellement la qualité du sommeil (un facteur important dans la difficulté de la prise en charge et l’activité nocturne des patients) [
31]. Il ne faut cependant pas s’exposer tout le temps à trop de lumière : une lumière excessive en soirée peut rendre le sommeil plus difficile à initier et moins profond en début de nuit [
32], et peut retarder notre horloge circadienne [1]. |
L’auteur déclare n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
|
1.
Czeisler
CA
,
Gooley
JJ
. Sleep and circadian rhythms in humans . Cold Spring Harb Symp Quant Biol.
2007; ; 72 : :579.–597. 2.
Stephenson
KM
,
Schroder
CM
,
Bertschy
G
,
Bourgin
P
. Complex interaction of circadian and non-circadian effects of light on mood: shedding new light on an old story . Sleep Med Rev.
2012; ; 16 : :445.–454. 3.
Chellappa
SL
,
Gordijn
MC
,
Cajochen
C
. Can light make us bright? Effects of light on cognition and sleep . Prog Brain Res.
2011; ; 190 : :119.–133. 4.
Vandewalle
G
,
Maquet
P
,
Dijk
DJ
. Light as a modulator of cognitive brain function . Trends Cogn Sci.
2009; ; 13 : :429.–438. 5.
Cajochen
C
,
Zeitzer
JM
,
Czeisler
CA
,
Dijk
DJ
. Dose-response relationship for light intensity and ocular and electroencephalographic correlates of human alertness . Behav Brain Res.
2000; ; 115 : :75.–83. 6.
Vandewalle
G
,
Schwartz
S
,
Grandjean
D
, et al.
Spectral quality of light modulates emotional brain responses in humans . Proc Natl Acad Sci USA.
2010; ; 107 : :19549.–19554. 7.
Vandewalle
G
,
Archer
SN
,
Wuillaume
C
, et al.
Effects of light on cognitive brain responses depend on circadian phase and sleep homeostasis . J Biol Rhythms.
2011; ; 26 : :249.–259. 8.
Vandewalle
G
,
Schmidt
C
,
Albouy
G
, et al.
Brain responses to violet, blue, green monochromatic light exposures in humans: prominent role of blue light, the brainstem . PLoS One.
2007; ; 2 : :e1247.. 9.
Perrin
F
,
Peigneux
P
,
Fuchs
S
, et al.
Nonvisual responses to light exposure in the human brain during the circadian night . Curr Biol.
2004; ; 14 : :1842.–1846. 10.
Jones
BE
. Arousal systems . Front Biosci.
2003; ; 8 : :s438.–s451. 11.
Chellappa
SL
,
Ly
JQ
,
Meyer
C
, et al.
Photic memory for executive brain responses . Proc Natl Acad Sci USA.
2014; ; 111 : :6087.–6091. 12.
Vandewalle
G
,
Balteau
E
,
Phillips
C
, et al.
Daytime light exposure dynamically enhances brain responses . Curr Biol.
2006; ; 16 : :1616.–1621. 13.
Vandewalle
G
,
Gais
S
,
Schabus
M
, et al.
Wavelength-dependent modulation of brain responses to a working memory task by daytime light exposure . Cereb Cortex.
2007; ; 17 : :2788.–2795. 14.
Koechlin
E
,
Hyafil
A
. Anterior prefrontal function and the limits of human decision-making . Science.
2007; ; 318 : :594.–598. 15.
Cajochen
C
,
Frey
S
,
Anders
D
, et al.
Evening exposure to a light-emitting diodes (LED)-backlit computer screen affects circadian physiology and cognitive performance . J Appl Physiol.
2011; ; 110 : :1432.–1438. 16.
Rahman
SA
,
Flynn-Evans
EE
,
Aeschbach
D
, et al.
Diurnal spectral sensitivity of the acute alerting effects of light . Sleep.
2014; ; 37 : :271.–281. 17.
Schmidt
TM
,
Chen
SK
,
Hattar
S
. Intrinsically photosensitive retinal ganglion cells: many subtypes, diverse functions . Trends Neurosci.
2011; ; 34 : :572.–580. 18.
Hatori
M
,
Panda
S
. The emerging roles of melanopsin in behavioral adaptation to light . Trends Mol Med.
2010; ; 16 : :435.–446. 19.
Vandewalle
G
,
Collignon
O
,
Hull
JT
, et al.
Blue light stimulates cognitive brain activity in visually blind individuals . J Cogn Neurosci.
2013; ; 25 : :2072.–2085. 20.
Mure
LS
,
Cornut
PL
,
Rieux
C
, et al.
Melanopsin bistability: a fly’s eye technology in the human retina . PLoS One.
2009; ; 4 : :e5991.. 21.
Cajochen
C
,
Munch
M
,
Kobialka
S
, et al.
High sensitivity of human melatonin, alertness, thermoregulation, and heart rate to short wavelength light . J Clin Endocrinol Metab.
2005; ; 90 : :1311.–1316. 22.
Lockley
SW
,
Evans
EE
,
Scheer
FAJL
, et al.
Short-wavelength sensitivity for the direct effects of light on alertness, vigilance, and the waking electroencephalogram in humans . Sleep.
2006; ; 29 : :161.–168. 23.
Dijk
DJ
,
Archer
SN
. PERIOD3, circadian phenotypes, and sleep homeostasis . Sleep Med Rev.
2010; ; 14 : :151.–160. 24.
Carrier
J
,
Viens
I
,
Poirier
G
, et al.
Sleep slow wave changes during the middle years of life . Eur J Neurosci.
2011; ; 33 : :758.–766. 25.
Sletten
TL
,
Revell
VL
,
Middleton
B
, et al.
Age-related changes in acute and phase-advancing responses to monochromatic light . J Biol Rhythms.
2009; ; 24 : :73.–84. 26.
Daneault
V
,
Vandewalle
G
,
Hébert
M
, et al.
Does pupil constriction under blue and green monochromatic light exposure change with age?
J Biol Rhythms.
2012; ; 27 : :257.–264. 27.
Najjar
RP
,
Chiquet
C
,
Teikari
P
, et al.
Aging of non-visual spectral sensitivity to light in humans: compensatory mechanisms?
PLoS One.
2014; ; 9 : :e85837.. 28.
Daneault
V
,
Hebert
M
,
Albouy
G
, et al.
Aging reduces the stimulating effect of blue light on cognitive brain functions . Sleep.
2014; ; 37 : :85.–96. 29.
Vandewalle
G
,
Hebert
M
,
Beaulieu
C
, et al.
Abnormal hypothalamic response to light in seasonal affective disorder . Biol Psychiatry.
2011; ; 70 : :954.–961. 30.
Viola
AU
,
James
LM
,
Schlangen
LJ
,
Dijk
DJ
. Blue-enriched white light in the workplace improves self-reported alertness, performance and sleep quality . Scand J Work Environ Health.
2008; ; 34 : :297.–306. 31.
Riemersma-van der Lek
RF
,
Swaab
DF
,
Twisk
J
, et al.
Effect of bright light and melatonin on cognitive and noncognitive function in elderly residents of group care facilities: a randomized controlled trial . JAMA.
2008; ; 299 : :2642.–2655. 32.
Chellappa
SL
,
Steiner
R
,
Oelhafen
P
, et al.
Acute exposure to evening blue-enriched light impacts on human sleep . J Sleep Res.
2013; ; 22 : :573.–580. 33.
Dijk
D
,
Archer
SN
. Light, sleep, circadian rhythms: together again . PLoS Biol.
2009; ; 7 : :e1000145.. 34.
Vandewalle
G
,
Dijk
DJ
. Neuroimaging the effects of light on non-visual brain functions . In :
Nofzinger
E
,
Maquet
P
,
Thorpy
M
, eds. Neuroimaging of sleep and sleep disorders . Cambridge (UK): : Cambridge University Press; , 2013 : :171.–178. 35.
Duez
H
,
Sebti
Y
,
Staels
B
. Horloges circadiennes et métabolisme : intégration des signaux métaboliques et environnementaux . Med Sci (Paris).
2013; ; 29 : :772.–777. 36.
Volle
E
,
Levy
R
. Rôle du cortex préfrontal dans l’adaptation comportementale chez l’homme . Med Sci (Paris).
2014; ; 30 : :179.–185. 37.
Gagne
AM
,
Bouchard
G
,
Tremblay
P
, et al.
Quand la saison devient synonyme de dépression . Med Sci (Paris).
2010; ; 26 : :79.–82. |






